

The Mre11/Rad50/Nbs1 complex functions in resection-based DNA end joining in Xenopus laevis
- PMID: 19892829
- PMCID: PMC2811014
- DOI: 10.1093/nar/gkp905
The Mre11/Rad50/Nbs1 complex functions in resection-based DNA end joining in Xenopus laevis
Abstract
The repair of DNA double-strand breaks (DSBs) is essential to maintain genomic integrity. In higher eukaryotes, DNA DSBs are predominantly repaired by non-homologous end joining (NHEJ), but DNA ends can also be joined by an alternative error-prone mechanism termed microhomology-mediated end joining (MMEJ). In MMEJ, the repair of DNA breaks is mediated by annealing at regions of microhomology and is always associated with deletions at the break site. In budding yeast, the Mre11/Rad5/Xrs2 complex has been demonstrated to play a role in both classical NHEJ and MMEJ, but the involvement of the analogous MRE11/RAD50/NBS1 (MRN) complex in end joining in higher eukaryotes is less certain. Here we demonstrate that in Xenopus laevis egg extracts, the MRN complex is not required for classical DNA-PK-dependent NHEJ. However, the XMRN complex is necessary for resection-based end joining of mismatched DNA ends. This XMRN-dependent end joining process is independent of the core NHEJ components Ku70 and DNA-PK, occurs with delayed kinetics relative to classical NHEJ and brings about repair at sites of microhomology. These data indicate a role for the X. laevis MRN complex in MMEJ.
Figures
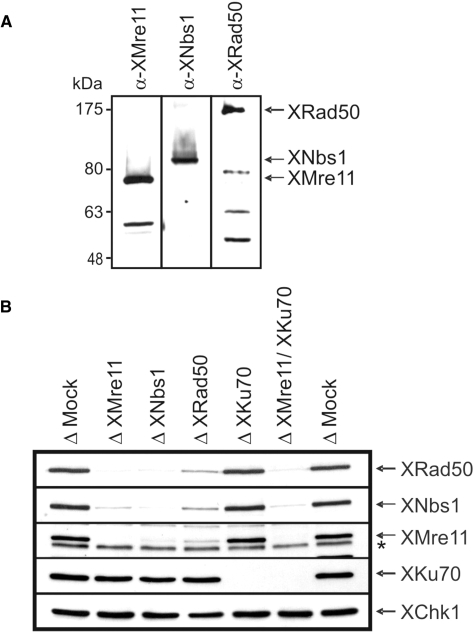
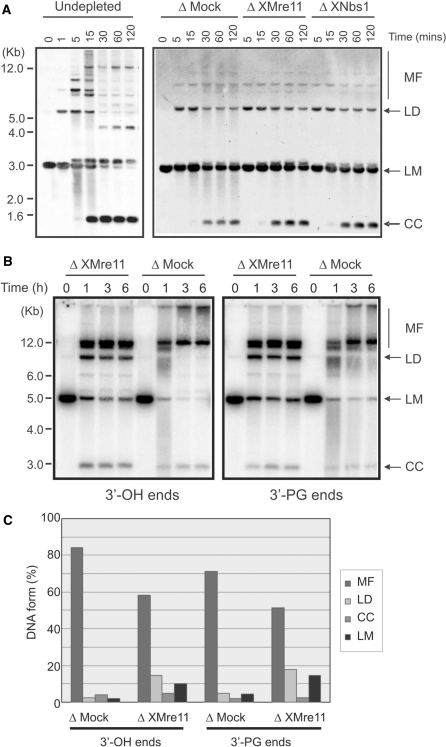
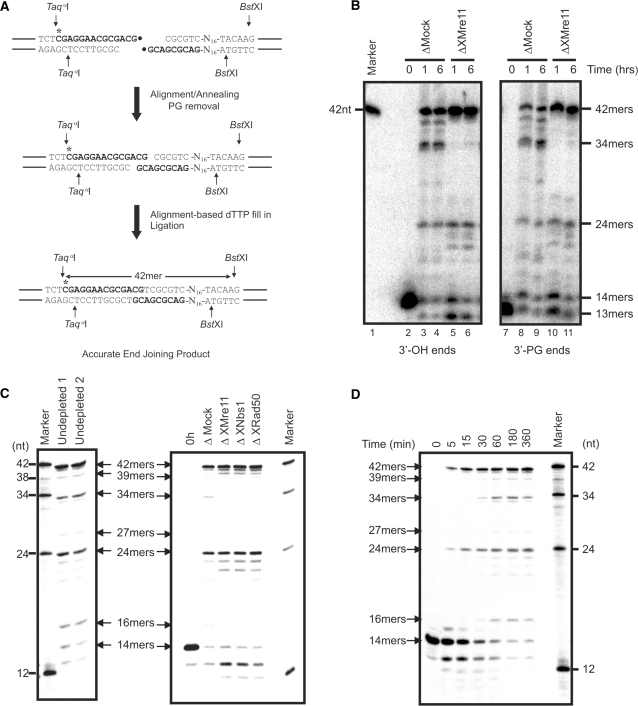
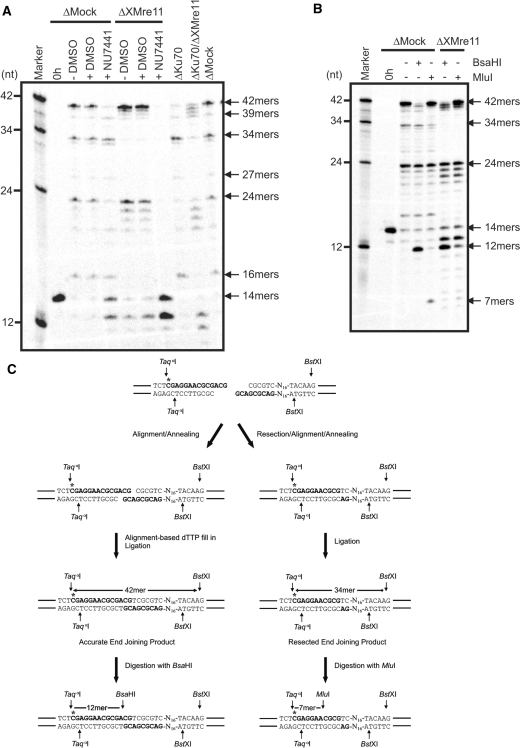
References
-
- Khanna KK, Jackson SP. DNA double-strand breaks: signaling, repair and the cancer connection. Nat. Genet. 2001;27:247–254. - PubMed
-
- van Gent DC, Hoeijmakers JH, Kanaar R. Chromosomal stability and the DNA double-stranded break connection. Nat. Rev. 2001;2:196–206. - PubMed
-
- Jazayeri A, Falck J, Lukas C, Bartek J, Smith GC, Lukas J, Jackson SP. ATM- and cell cycle-dependent regulation of ATR in response to DNA double-strand breaks. Nat. Cell. Biol. 2006;8:37–45. - PubMed
-
- West SC. Molecular views of recombination proteins and their control. Nat. Rev. Mol. Cell. Biol. 2003;4:435–445. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
