

Origin of cardiac fibroblasts and the role of periostin
- PMID: 19893021
- PMCID: PMC2786053
- DOI: 10.1161/CIRCRESAHA.109.201400
Origin of cardiac fibroblasts and the role of periostin
Abstract
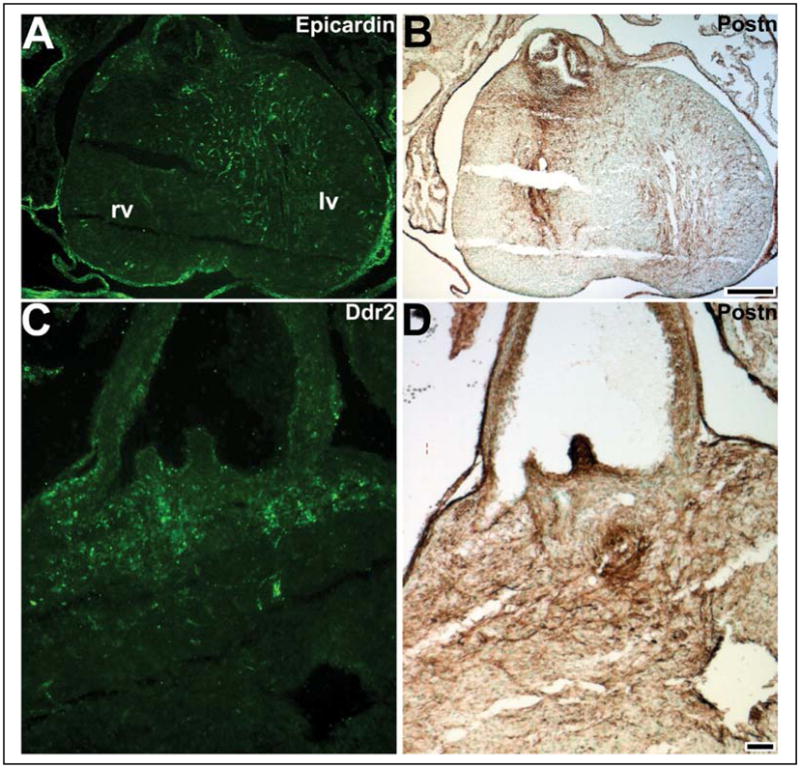
Cardiac fibroblasts are the most populous nonmyocyte cell type within the mature heart and are required for extracellular matrix synthesis and deposition, generation of the cardiac skeleton, and to electrically insulate the atria from the ventricles. Significantly, cardiac fibroblasts have also been shown to play an important role in cardiomyocyte growth and expansion of the ventricular chambers during heart development. Although there are currently no cardiac fibroblast-restricted molecular markers, it is generally envisaged that the majority of the cardiac fibroblasts are derived from the proepicardium via epithelial-to-mesenchymal transformation. However, still relatively little is known about when and where the cardiac fibroblasts cells are generated, the lineage of each cell, and how cardiac fibroblasts move to reside in their final position throughout all four cardiac chambers. In this review, we summarize the present understanding regarding the function of Periostin, a useful marker of the noncardiomyocyte lineages, and its role during cardiac morphogenesis. Characterization of the cardiac fibroblast lineage and identification of the signals that maintain, expand and regulate their differentiation will be required to improve our understanding of cardiac function in both normal and pathophysiological states.
Figures





References
-
- Slater M. Dynamic interactions of the extracellular matrix. Histol Histopathol. 1996;11:175–180. - PubMed
-
- Eghbali M, Czaja MJ, Zeydel M, Weiner FR, Zern MA, Seifter S, Blumenfeld OO. Collagen chain mRNAs in isolated heart cells from young and adult rats. J Mol Cell Cardiol. 1988;20:267–276. - PubMed
-
- Cleutjens JP, Kandala JC, Guarda E, Guntaka RV, Weber KT. Regulation of collagen degradation in the rat myocardium after infarction. J Mol Cell Cardiol. 1995;27:1281–1292. - PubMed
-
- Peterson JT, Li H, Dillon L, Bryant JW. Evolution of matrix metalloprotease and tissue inhibitor expression during heart failure progression in the infarcted rat. Cardiovasc Res. 2000;46:307–315. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
