

NF45 functions as an IRES trans-acting factor that is required for translation of cIAP1 during the unfolded protein response
- PMID: 19893574
- PMCID: PMC5017871
- DOI: 10.1038/cdd.2009.164
NF45 functions as an IRES trans-acting factor that is required for translation of cIAP1 during the unfolded protein response
Abstract
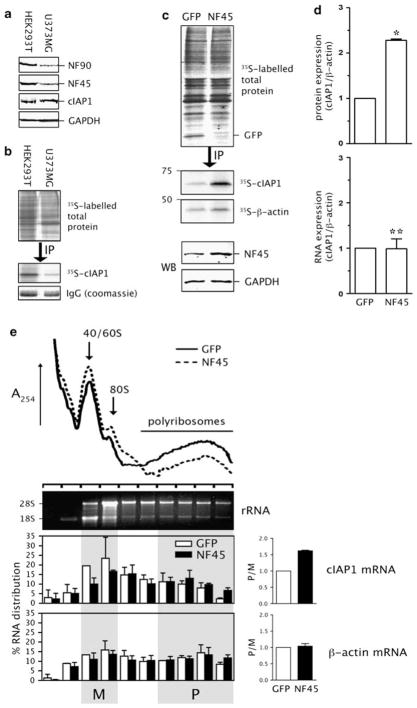
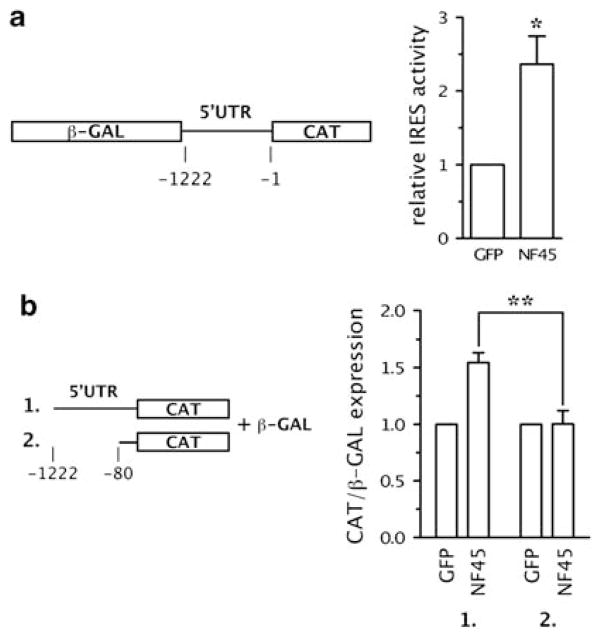
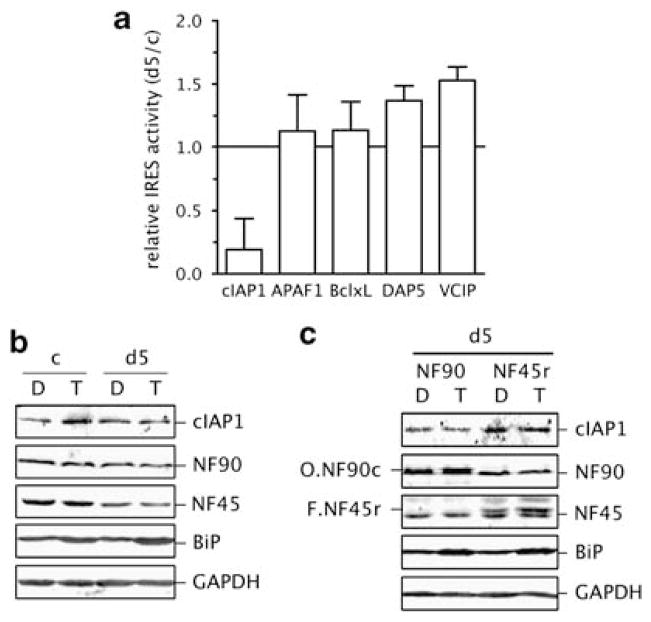
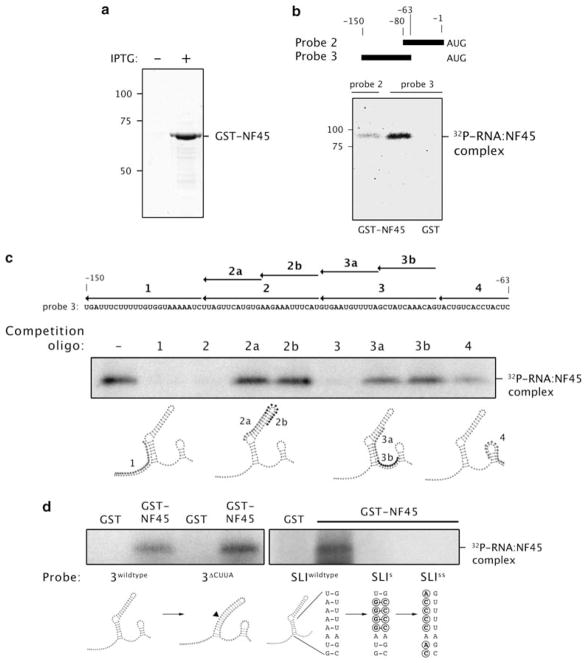
Expression of the cellular inhibitor of apoptosis protein 1 (cIAP1) is unexpectedly repressed at the level of translation under normal physiological conditions in many cell lines. We have previously shown that the 5' untranslated region of cIAP1 mRNA contains a stress-inducible internal ribosome entry site (IRES) that governs expression of cIAP1 protein. Although inactive in unstressed cells, the IRES supports cap-independent translation of cIAP1 in response to endoplasmic reticulum stress. To gain an insight into the mechanism of cIAP1 IRES function, we empirically derived the minimal free energy secondary structure of the cIAP1 IRES using enzymatic cleavage mapping. We subsequently used RNA affinity chromatography to identify several cellular proteins, including nuclear factor 45 (NF45) as cIAP1 IRES binding proteins. In this report we show that NF45 is a novel RNA binding protein that enhances IRES-dependent translation of endogenous cIAP1. Further, we show that NF45 is required for IRES-mediated induction of cIAP1 protein during the unfolded protein response. The data presented are consistent with a model in which translation of cIAP1 is governed, at least in part, by NF45, a novel cellular IRES trans-acting factor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Qin X, Sarnow P. Preferential translation of internal ribosome entry site-containing mRNAs during the mitotic cycle in mammalian cells. J Biol Chem. 2004;279:13721–13728. - PubMed
-
- Gerlitz G, Jagus R, Elroy-Stein O. Phosphorylation of initiation factor-2 alpha is required for activation of internal translation initiation during cell differentiation. Eur J Biochem. 2002;269:2810–2819. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
