

Integration of a phosphatase cascade with the mitogen-activated protein kinase pathway provides for a novel signal processing function
- PMID: 19897477
- PMCID: PMC2801257
- DOI: 10.1074/jbc.M109.055863
Integration of a phosphatase cascade with the mitogen-activated protein kinase pathway provides for a novel signal processing function
Abstract
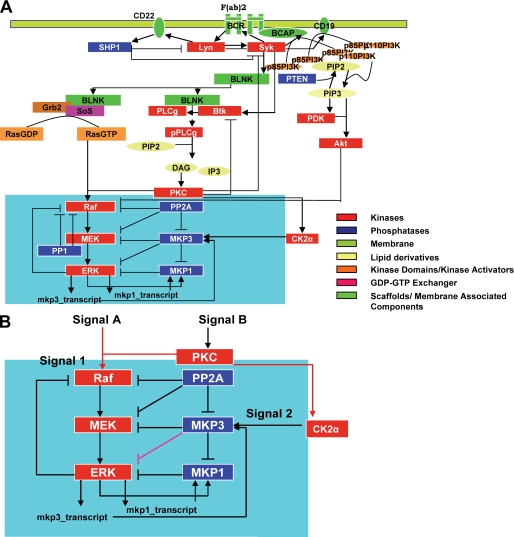
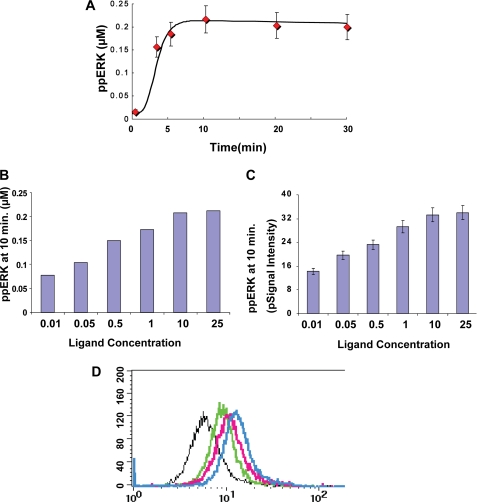
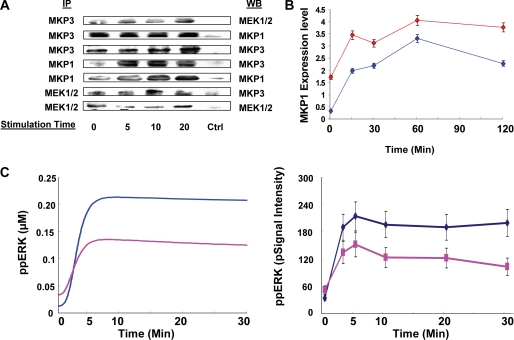
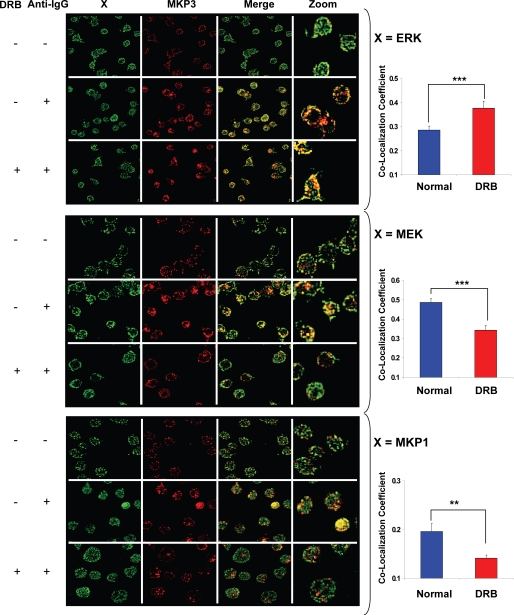
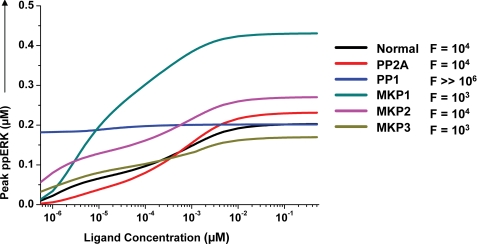
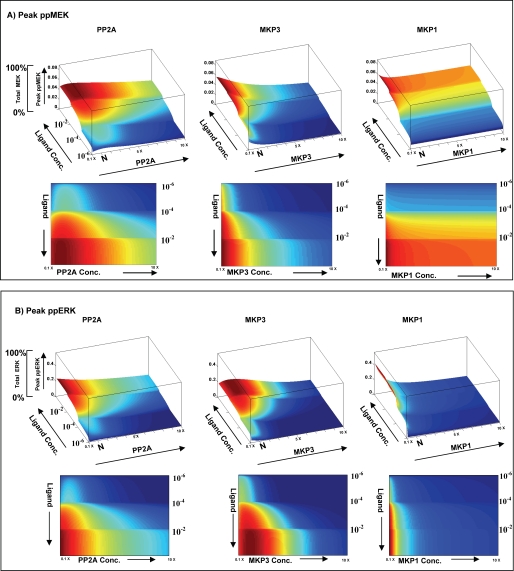
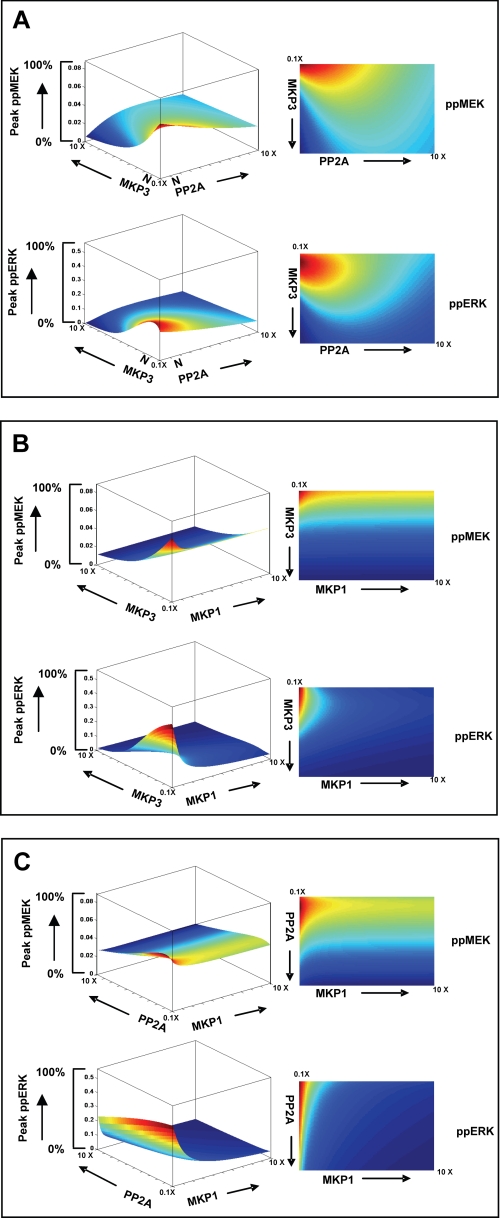
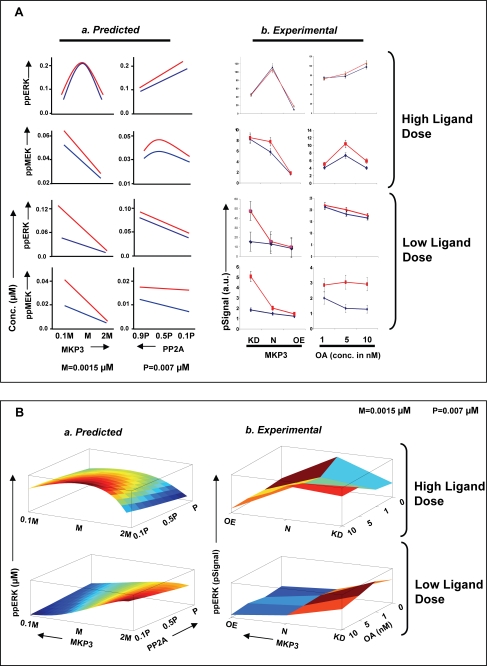
We mathematically modeled the receptor-dependent mitogen-activated protein kinase (MAPK) signaling by incorporating the regulation through cellular phosphatases. Activation induced the alignment of a phosphatase cascade in parallel with the MAPK pathway. A novel regulatory motif was, thus, generated, providing for the combinatorial control of each MAPK intermediate. This ensured a non-linear mode of signal transmission with the output being shaped by the balance between the strength of input signal and the activity gradient along the phosphatase axis. Shifts in this balance yielded modulations in topology of the motif, thereby expanding the repertoire of output responses. Thus, we identify an added dimension to signal processing wherein the output response to an external stimulus is additionally filtered through indicators that define the phenotypic status of the cell.
Figures
References
-
- Cobb M. H. (1999) Prog. Biophys. Mol. Biol. 71, 479–500 - PubMed
-
- Widmann C., Gibson S., Jarpe M. B., Johnson G. L. (1999) Physiol. Rev. 79, 143–180 - PubMed
-
- Chang L., Karin M. (2001) Nature 410, 37–40 - PubMed
-
- Tan P. B., Kim S. K. (1999) Trends Genet 15, 145–149 - PubMed
-
- Ebisuya M., Kondoh K., Nishida E. (2005) J. Cell Sci. 118, 2997–3002 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
