

A genomic island defines subspecies-specific virulence features of the host-adapted pathogen Campylobacter fetus subsp. venerealis
- PMID: 19897645
- PMCID: PMC2805309
- DOI: 10.1128/JB.00803-09
A genomic island defines subspecies-specific virulence features of the host-adapted pathogen Campylobacter fetus subsp. venerealis
Abstract
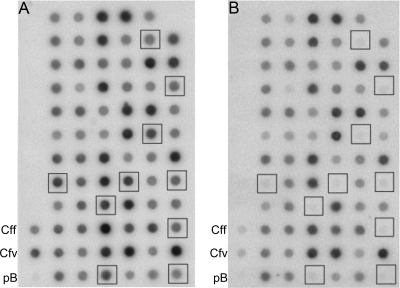
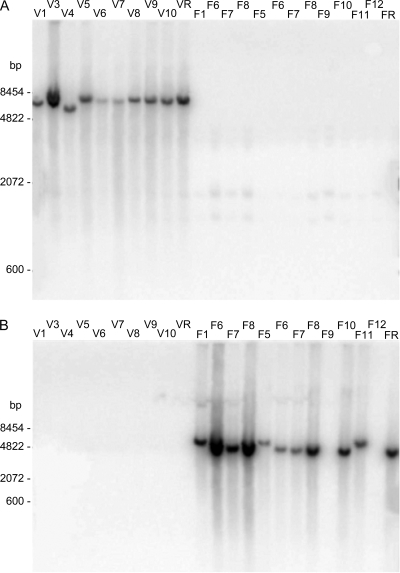
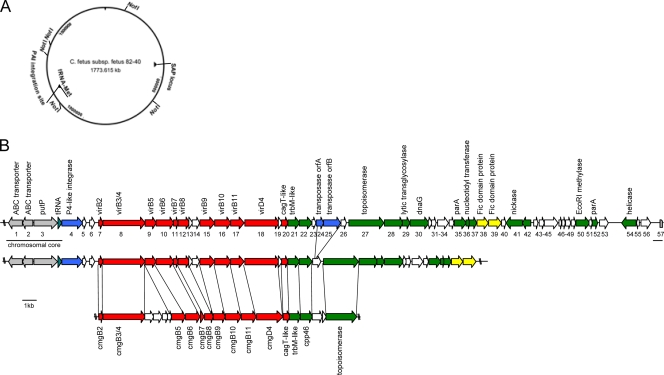
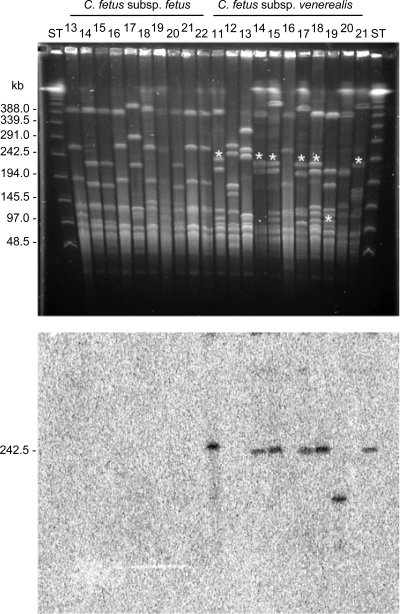
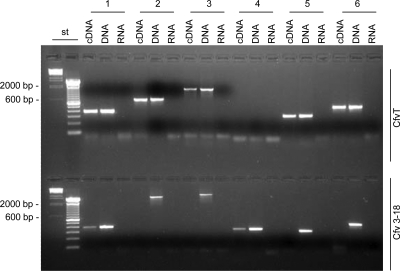
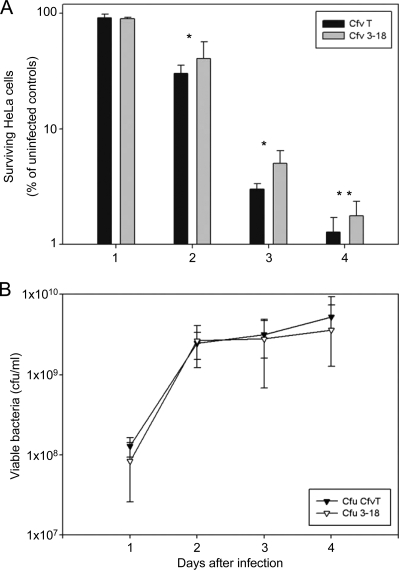
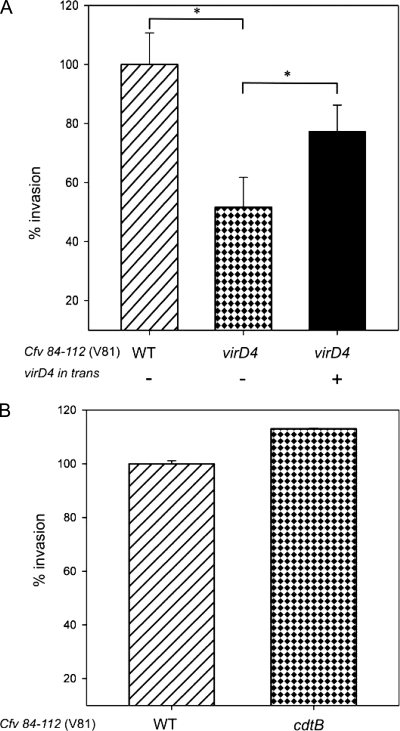
The pathogen Campylobacter fetus comprises two subspecies, C. fetus subsp. fetus and C. fetus subsp. venerealis. Although these taxa are highly related on the genome level, they are adapted to distinct hosts and tissues. C. fetus subsp. fetus infects a diversity of hosts, including humans, and colonizes the gastrointestinal tract. In contrast, C. fetus subsp. venerealis is largely restricted to the bovine genital tract, causing epidemic abortion in these animals. In light of their close genetic relatedness, the specific niche preferences make the C. fetus subspecies an ideal model system to investigate the molecular basis of host adaptation. In this study, a subtractive-hybridization approach was applied to the genomes of the subspecies to identify different genes potentially underlying this specificity. The comparison revealed a genomic island uniquely present in C. fetus subsp. venerealis that harbors several genes indicative of horizontal transfer and that encodes the core components necessary for bacterial type IV secretion. Macromolecular transporters of this type deliver effector molecules to host cells, thereby contributing to virulence in various pathogens. Mutational inactivation of the putative secretion system confirmed its involvement in the pathogenicity of C. fetus subsp. venerealis.
Figures
References
-
- Anonymous. 1996. OIE manual of standards for diagnostic tests and vaccines, 3rd ed., p. 256-266. Office International des Epizooties, Paris, France.
-
- Asakura, M., W. Samosornsuk, M. Taguchi, K. Kobayashi, N. Misawa, M. Kusumoto, K. Nishimura, A. Matsuhisa, and S. Yamasaki. 2007. Comparative analysis of cytolethal distending toxin (cdt) genes among Campylobacter jejuni, C. coli and C. fetus strains. Microb. Pathog. 42:174-183. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. A. Smith, J. G. Seidman, and K. Struhl. 1987. Current protocols in molecular biology. John Wiley & Sons, New York, NY.
-
- Baar, C., M. Eppinger, G. Raddatz, J. Simon, C. Lanz, O. Klimmek, R. Nandakumar, R. Gross, A. Rosinus, H. Keller, P. Jagtap, B. Linke, F. Meyer, H. Lederer, and S. C. Schuster. 2003. Complete genome sequence and analysis of Wolinella succinogenes. Proc. Natl. Acad. Sci. U. S. A. 100:11690-11695. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
