

Strain-dependent activation of NF-kappaB in the airway epithelium and its role in allergic airway inflammation
- PMID: 19897746
- PMCID: PMC2806198
- DOI: 10.1152/ajplung.00037.2009
Strain-dependent activation of NF-kappaB in the airway epithelium and its role in allergic airway inflammation
Abstract
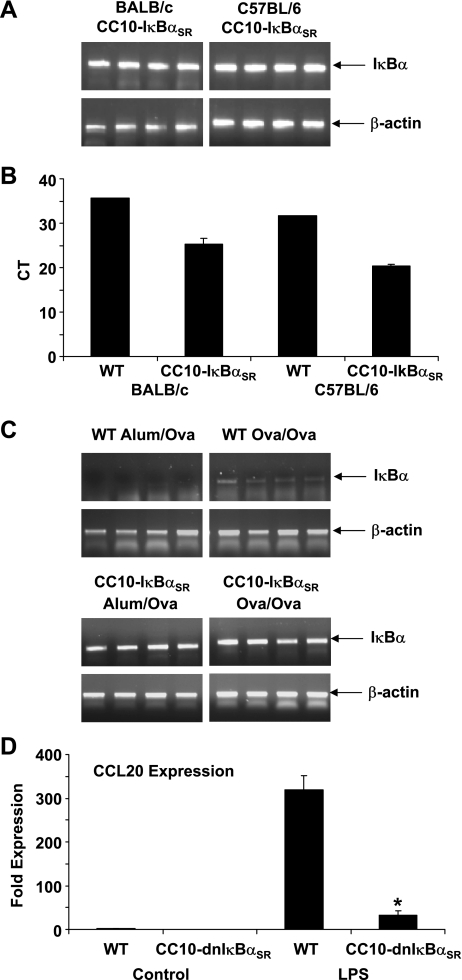
NF-kappaB activation in the airway epithelium has been established as a critical pathway in ovalbumin (Ova)-induced airway inflammation in BALB/c mice (Poynter ME, Cloots R, van Woerkom T, Butnor KJ, Vacek P, Taatjes DJ, Irvin CG, Janssen-Heininger YM. J Immunol 173: 7003-7009, 2004). BALB/c mice are susceptible to the development of allergic airway disease, whereas other strains of mice, such as C57BL/6, are considered more resistant. The goal of the present study was to determine the proximal signals required for NF-kappaB activation in the airway epithelium in allergic airway disease and to unravel whether these signals are strain-dependent. Our previous studies, conducted in the BALB/c mouse background, demonstrated that transgenic mice expressing a dominant-negative version of IkappaBalpha in the airway epithelium (CC10-IkappaBalpha(SR)) were protected from Ova-induced inflammation. In contrast to these earlier observations, we demonstrate here that CC10-IkappaBalpha(SR) transgenic mice on the C57BL/6 background were not protected from Ova-induced allergic airway inflammation. Consistent with this finding, Ova-induced nuclear localization of the RelA subunit of NF-kappaB was not observed in C57BL/6 mice, in contrast to the marked nuclear presence of RelA in BALB/c mice. Evaluation of cytokine profiles in bronchoalveolar lavage demonstrated elevated expression of TNF-alpha in BALB/c mice compared with C57BL/6 mice after an acute challenge with Ova. Finally, neutralization of TNF-alpha by a blocking antibody prevented nuclear localization of RelA in BALB/c mice after Ova challenge. These data suggest that the mechanism of response of the airway epithelium of immunized C57BL/6 mice to antigen challenge is fundamentally different from that of immunized BALB/c mice and highlight the potential importance of TNF-alpha in regulating epithelial NF-kappaB activation in allergic airway disease.
Figures





Similar articles
-
Distinct functions of airway epithelial nuclear factor-kappaB activity regulate nitrogen dioxide-induced acute lung injury.Am J Respir Cell Mol Biol. 2010 Oct;43(4):443-51. doi: 10.1165/rcmb.2008-0416OC. Epub 2009 Nov 9. Am J Respir Cell Mol Biol. 2010. PMID: 19901348 Free PMC article.
-
Epithelial NF-κB orchestrates house dust mite-induced airway inflammation, hyperresponsiveness, and fibrotic remodeling.J Immunol. 2013 Dec 15;191(12):5811-21. doi: 10.4049/jimmunol.1301329. Epub 2013 Nov 13. J Immunol. 2013. PMID: 24227776 Free PMC article.
-
A prominent role for airway epithelial NF-kappa B activation in lipopolysaccharide-induced airway inflammation.J Immunol. 2003 Jun 15;170(12):6257-65. doi: 10.4049/jimmunol.170.12.6257. J Immunol. 2003. PMID: 12794158
-
NF-kappa B activation in airways modulates allergic inflammation but not hyperresponsiveness.J Immunol. 2004 Dec 1;173(11):7003-9. doi: 10.4049/jimmunol.173.11.7003. J Immunol. 2004. PMID: 15557197 Free PMC article.
-
Airway epithelial regulation of allergic sensitization in asthma.Pulm Pharmacol Ther. 2012 Dec;25(6):438-46. doi: 10.1016/j.pupt.2012.04.005. Epub 2012 May 9. Pulm Pharmacol Ther. 2012. PMID: 22579987 Free PMC article. Review.
Cited by
-
Characterization of a novel high-dose ovalbumin-induced murine model of allergic sinonasal inflammation.Int Forum Allergy Rhinol. 2016 Sep;6(9):964-72. doi: 10.1002/alr.21768. Epub 2016 Apr 5. Int Forum Allergy Rhinol. 2016. PMID: 27060366 Free PMC article.
-
The receptor for advanced glycation end products is a central mediator of asthma pathogenesis.Am J Pathol. 2012 Oct;181(4):1215-25. doi: 10.1016/j.ajpath.2012.06.031. Epub 2012 Aug 11. Am J Pathol. 2012. PMID: 22889845 Free PMC article.
-
Protease-activated receptor-2 activation contributes to house dust mite-induced IgE responses in mice.PLoS One. 2014 Mar 20;9(3):e91206. doi: 10.1371/journal.pone.0091206. eCollection 2014. PLoS One. 2014. PMID: 24651123 Free PMC article.
-
IL-22 is essential for lung epithelial repair following influenza infection.Am J Pathol. 2013 Apr;182(4):1286-96. doi: 10.1016/j.ajpath.2012.12.007. Epub 2013 Mar 11. Am J Pathol. 2013. PMID: 23490254 Free PMC article.
-
Long-chain acyl-CoA dehydrogenase deficiency as a cause of pulmonary surfactant dysfunction.J Biol Chem. 2014 Apr 11;289(15):10668-10679. doi: 10.1074/jbc.M113.540260. Epub 2014 Mar 3. J Biol Chem. 2014. PMID: 24591516 Free PMC article.
References
-
- Aoki T, Hirota T, Tamari M, Ichikawa K, Takeda K, Arinami T, Shibasaki M, Noguchi E. An association between asthma and TNF-308G/A polymorphism: meta-analysis. J Hum Genet 51: 677–685, 2006 - PubMed
-
- Bates JH, Irvin CG. Measuring lung function in mice: the phenotyping uncertainty principle. J Appl Physiol 94: 1297–1306, 2003 - PubMed
-
- Berry MA, Hargadon B, Shelley M, Parker D, Shaw DE, Green RH, Bradding P, Brightling CE, Wardlaw AJ, Pavord ID. Evidence of a role of tumor necrosis factor-α in refractory asthma. N Engl J Med 354: 697–708, 2006 - PubMed
-
- Bilolikar H, Nam AR, Rosenthal M, Davies JC, Henderson DC, Balfour-Lynn IM. Tumour necrosis factor gene polymorphisms and childhood wheezing. Eur Respir J 26: 637–646, 2005 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous