

Review
doi: 10.1038/nrg2697.
Epub 2009 Nov 10.
Synthetic biology: understanding biological design from synthetic circuits
Affiliations
- PMID: 19898500
- PMCID: PMC3138802
- DOI: 10.1038/nrg2697
Item in Clipboard
Review
Synthetic biology: understanding biological design from synthetic circuits
Nat Rev Genet.
2009 Dec.
Abstract
An important aim of synthetic biology is to uncover the design principles of natural biological systems through the rational design of gene and protein circuits. Here, we highlight how the process of engineering biological systems - from synthetic promoters to the control of cell-cell interactions - has contributed to our understanding of how endogenous systems are put together and function. Synthetic biological devices allow us to grasp intuitively the ranges of behaviour generated by simple biological circuits, such as linear cascades and interlocking feedback loops, as well as to exert control over natural processes, such as gene expression and population dynamics.
Figures
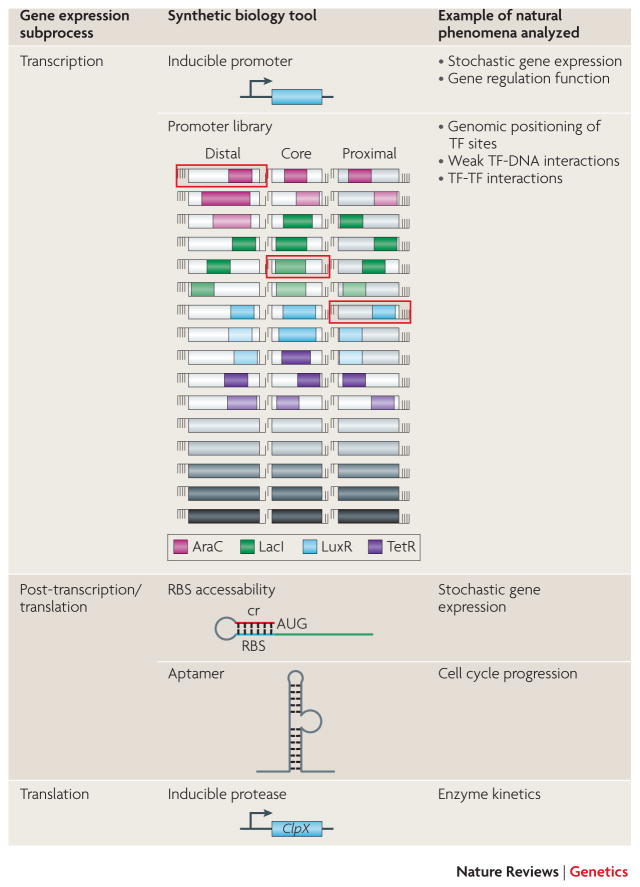
The diagram shows the transcriptional and post-transcriptional processes in gene expression that can be manipulated by synthetic biology tools, and some example applications. TF, transcription factor; RBS, ribosome binding site. Promoter library diagram from Cox et al. Cox RS 3rd, Surette MG, Elowitz MB. Programming gene expression with combinatorial promoters. Mol. Syst. Bio. 3: 145 (2007) RBS accessibility diagram from Isaacs et al. Isaacs FJ, Dwyer DJ, Ding C, Pervouchine DD, Cantor CR, Collins JJ. Engineered riboregulators enable post-transcriptional control of gene expression. Nat Biotech 22, 841–847 (2004) Aptamer diagram from Grate and Wilson Grate D, Wilson C. Inducible regulation of the S. cerevisiae cell cycle mediated by an RNA aptamer-ligand complex. Bioorg Med Chem 9: 2565–2570 (2001)
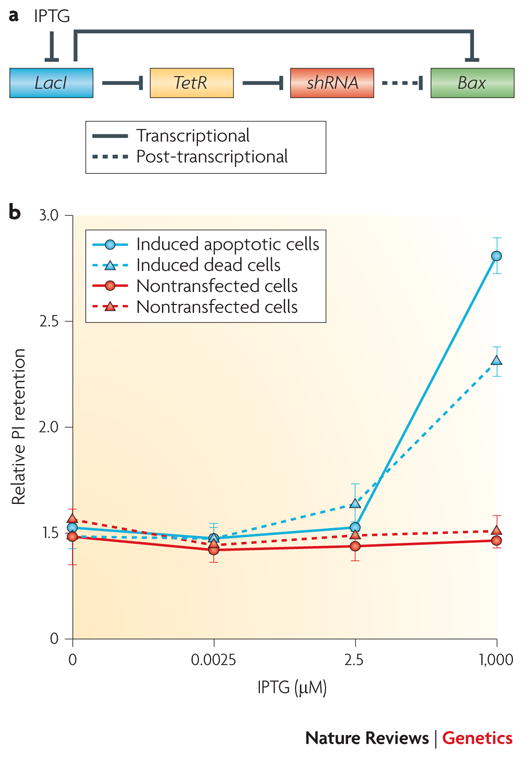
A) Deans et al. created a genetic switch whose state is read out by a GFP reporter or a gene of interest; here, the gene of interest that we focus on is Bax, a pro-apoptotic gene. Bax is under the transcriptional control of the Lac repressor (LacI) and under the translational control of a short hairpin (sh)RNA, which itself is under transcriptional control of the TetR repressor. In the “OFF” state, LacI inhibits transcription of Bax. Additionally, LacI inhibits transcription of the TetR repressor; this allows the transcription of the shRNA, which goes on to inhibit translation of Bax by cleaving its mRNA. The result of this dual-layered repression is the creation of a truly off “OFF” state; whereas in the initial characterization each mode of repression alone was able to reduce reporter levels by about 80%, leaving a BASAL EXPRESSION of 20%, the combination resulted in greater than 99% repression. The circuit can then be tunably activated by adding varying amounts of IPTG, which blocks the effects of LacI. B) The fraction of cells that undergo apoptosis is determined by Bax expression levels. Data obtained by tuning Bax with IPTG, as described above, offer some tantalizing clues into the fundamental molecular biology underlying the apoptosis pathway. In particular the data are consistent with the idea that the decision to undergo apoptosis (assessed by retention of propidium iodide (PI) dye relative to PI retention due to the transfection protocol alone) is determined by reaching a threshold level of Bax. Although the Bax threshold data are not conclusive, the result demonstrates the power of a technique that allows one to rationally tune the level of any gene of interest and examine the consequences. Panel B is reproduced with permission from Figure 6b of Deans TL, Cantor CR, Collins JJ. A tunable genetic switch based on RNAi and repressor proteins for regulating gene expression in mammalian cells. Cell 130, 363–372 (2007)
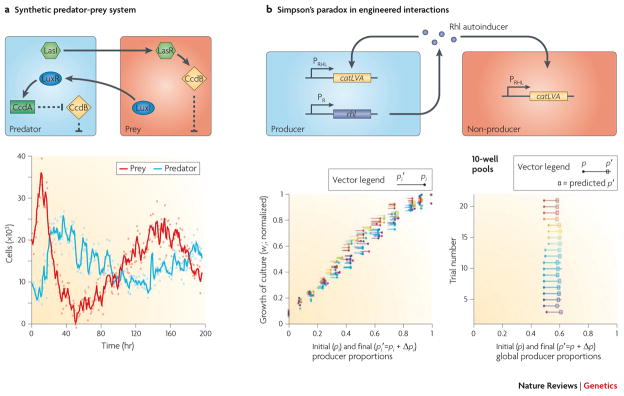
Studies of ecology and evolution are often dependent on carefully characterizing the interactions of different organisms. In a natural setting, however, such data collection often proves to be noisy at best and impossible at worst. At the same time, mathematical models in theoretical ecology and evolutionary biology are among the most sophisticated in all of the life sciences. Laboratory-scale experiments on cellular interactions could quantitatively test some of the remarkable predictions and open the way to new theory. A) Among the most elementary interactions in nature is the predator-prey interaction. The prey in this case produces the quorum sensing pathway protein LuxI, which is engineered to drive a transcriptional cascade in the predator that produces CcdA, which inhibits the DNA replication inhibitor CcdB thereby allowing the predators to replicate. Meanwhile the predator produces the quorum sensing pathway protein LasI which activates CcdB in a LasR-dependent manner in the prey. CcdB expression in the prey prevents it from replicating. The cyclic dynamic is very similar in style to genetic oscillators: high levels of prey leads to low levels of CcdB and thus high levels of predator; high levels of predator leads to high levels of CcdB and thus low levels of prey, which subsequently leads to high levels of CcdB in predators, etc. As shown in Balagadde et al., predator-prey interactions can thus result in LIMIT CYCLE OSCILLATIONS about an unstable fixed point of the dynamics, most commonly studied in the framework of the Lotka-Volterra model. B) Simpson’s paradox is a statistical phenomenon that captures the fact that even if the producer of a common good grows at a slower rate in all given subpopulations than a nonproducer, it can nevertheless make up an increasing fraction of the population as a whole. While Simpson’s paradox usually arises as a result of misinterpretation of data, natural populations can in fact display heterogeneities in sample size that often underlie the paradox. The particular implementation in Chuang et al. casts bacteria that generate the autoinducer Rhl as the producer. Both producer and non producers use this Rhl that is rewired to activate synthesis of a chloramphenicol resistance gene catLVA. As shown in the middle panel, in each subpopulation the fraction of producers decreases, but as the bottom panel shows in the global population the fraction of producers actually increases, thus satisfying Simpson’s paradox. (A, lower portion) is reproduced with permission from Balagadde FK, Song H, Ozaki J, Collins CH, Barnet M, Arnold FH, Quake SR, You L. A synthetic Escherichia coli predator-prey ecosystem. Mol Syst Biol 4: 187 (2008) © Macmillan Publishing Ltd. Panel (B) lower subpanels are reproduced from Figure 2a and 2b[add source of top panel] of Chuang et al. Chuang JS, Rivoire O, Leibler S. Simpson’s paradox in a synthetic microbial system. Science 323(5911): 272–275 (2009)
References
-
- Mattick JS. RNA regulation: a new genetics? Nat Rev Genet. 2004;5:316–323. - PubMed
-
- Davidson EA. The regulatory genome: gene regulatory networks in development and evolution. Academic Press; 2006.
-
- Bridgham JT, Carroll SM, Thornton JW. Evolution of hormone-receptor complexity by molecular exploitation. Science. 2006;312:97–101. Uses rational synthesis of a resurrected glucocorticoid receptor to explore the evolution of the receptor ligand–pair. - PubMed
-
- Rapp M, Seppala S, Granseth E, von Heijne G. Emulating membrane protein evolution by rational design. Science. 2007;315:1282–1284. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
