

Glycosylation regulates prestin cellular activity
- PMID: 19898896
- PMCID: PMC2820205
- DOI: 10.1007/s10162-009-0196-5
Glycosylation regulates prestin cellular activity
Abstract
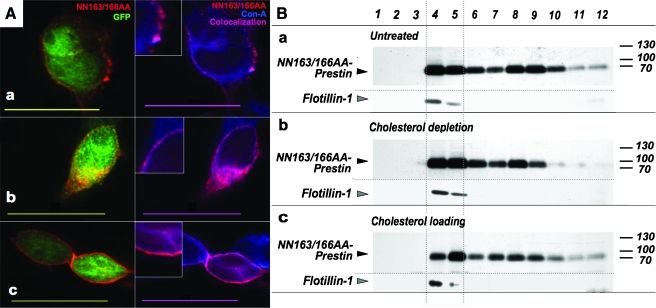
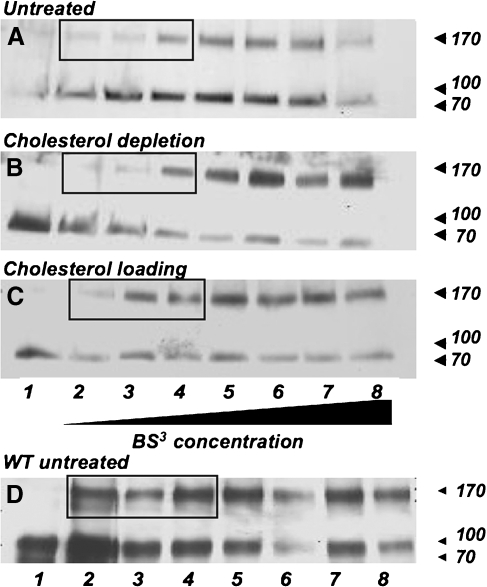
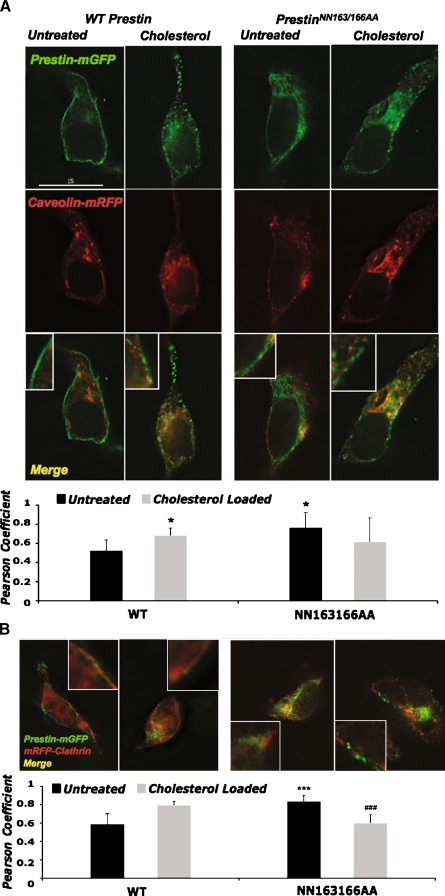
Glycosylation is a common post-translational modification of proteins and is implicated in a variety of cellular functions including protein folding, degradation, sorting and trafficking, and membrane protein recycling. The membrane protein prestin is an essential component of the membrane-based motor driving electromotility changes (electromotility) in the outer hair cell (OHC), a central process in auditory transduction. Prestin was earlier identified to possess two N-glycosylation sites (N163, N166) that, when mutated, marginally affect prestin nonlinear capacitance (NLC) function in cultured cells. Here, we show that the double mutant prestin(NN163/166AA) is not glycosylated and shows the expected NLC properties in the untreated and cholesterol-depleted HEK 293 cell model. In addition, unlike WT prestin that readily forms oligomers, prestin(NN163/166AA) is enriched as monomers and more mobile in the plasma membrane, suggesting that oligomerization of prestin is dependent on glycosylation but is not essential for the generation of NLC in HEK 293 cells. However, in the presence of increased membrane cholesterol, unlike the hyperpolarizing shift in NLC seen with WT prestin, cells expressing prestin(NN163/166AA) exhibit a linear capacitance function. In an attempt to explain this finding, we discovered that both WT prestin and prestin(NN163/166AA) participate in cholesterol-dependent cellular trafficking. In contrast to WT prestin, prestin(NN163/166AA) shows a significant cholesterol-dependent decrease in cell-surface expression, which may explain the loss of NLC function. Based on our observations, we conclude that glycosylation regulates self-association and cellular trafficking of prestin(NN163/166AA). These observations are the first to implicate a regulatory role for cellular trafficking and sorting in prestin function. We speculate that the cholesterol regulation of prestin occurs through localization to and internalization from membrane microdomains by clathrin- and caveolin-dependent mechanisms.
Figures



References
-
- Brownell WE. The piezoelectric hair cell. In: Eatock RA, editor. Vertebrate hair cells. New York: Springer; 2006. pp. 313–347.
-
- Brownell WE, Oghalai JS. Structural basis of outer hair cell motility or where's the motor? In: Lim D, editor. Cell and molecular biology of the ear. New York: Academic; 2000. pp. 69–83.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
