

Mapping of contextual modulation in the population response of primary visual cortex
- PMID: 19898958
- PMCID: PMC2837531
- DOI: 10.1007/s11571-009-9098-9
Mapping of contextual modulation in the population response of primary visual cortex
Abstract
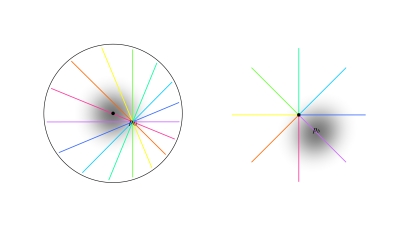
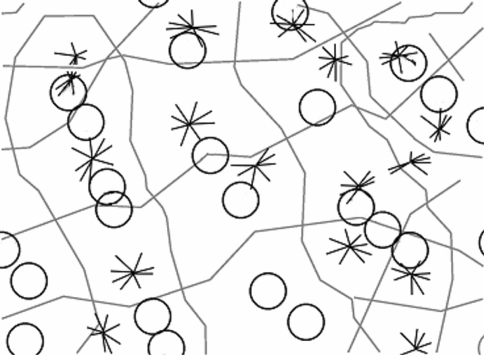
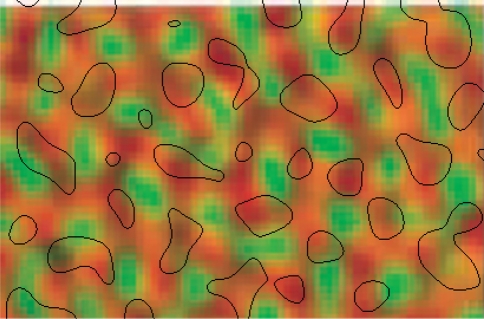
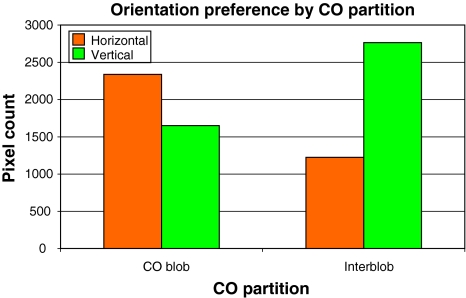
We review the evidence of long-range contextual modulation in V1. Populations of neurons in V1 are activated by a wide variety of stimuli outside of their classical receptive fields (RF), well beyond their surround region. These effects generally involve extra-RF features with an orientation component. The population mapping of orientation preferences to the upper layers of V1 is well understood, as far as the classical RF properties are concerned, and involves organization into pinwheel-like structures. We introduce a novel hypothesis regarding the organization of V1's contextual response. We show that RF and extra-RF orientation preferences are mapped in related ways. Orientation pinwheels are the foci of both types of features. The mapping of contextual features onto the orientation pinwheel has a form that recapitulates the organization of the visual field: an iso-orientation patch within the pinwheel also responds to extra-RF stimuli of the same orientation. We hypothesize that the same form of mapping applies to other stimulus properties that are mapped out in V1, such as colour and contrast selectivity. A specific consequence is that fovea-like properties will be mapped in a systematic way to orientation pinwheels. We review the evidence that cytochrome oxidase blobs comprise the foci of this contextual remapping for colour and low contrasts. Neurodynamics and motion in the visual field are argued to play an important role in the shaping and maintenance of this type of mapping in V1.
Figures
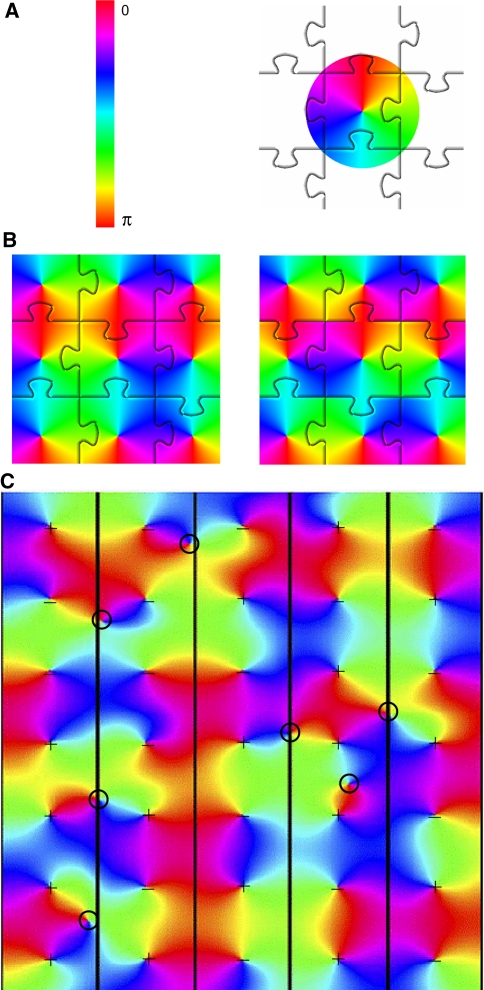

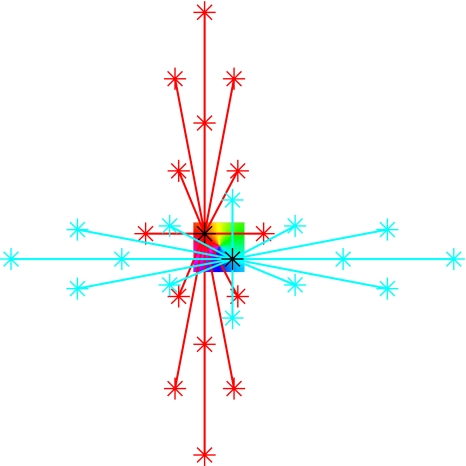
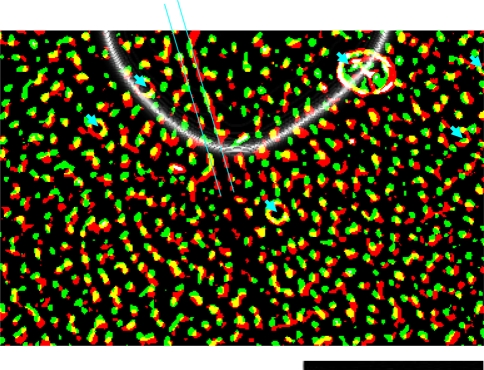
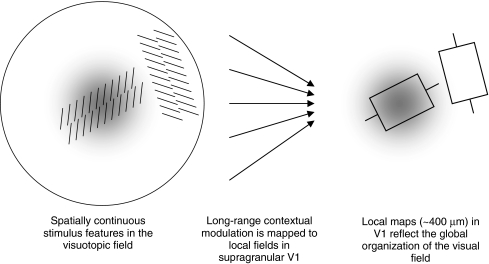


References
-
- Alexander DM, Bourke PD, Sheridan P, Konstandatos O, Wright JJ (1998) Emergent symmetry of local and global maps in the primary visual cortex: self-organization of orientation preference. Complexity International 6: http://www.complexity.org.au/ci/vol06/alexander/alexander.html
LinkOut - more resources
Full Text Sources
