

Plasma cell differentiation initiates a limited ER stress response by specifically suppressing the PERK-dependent branch of the unfolded protein response
- PMID: 19898960
- PMCID: PMC2866998
- DOI: 10.1007/s12192-009-0142-9
Plasma cell differentiation initiates a limited ER stress response by specifically suppressing the PERK-dependent branch of the unfolded protein response
Abstract
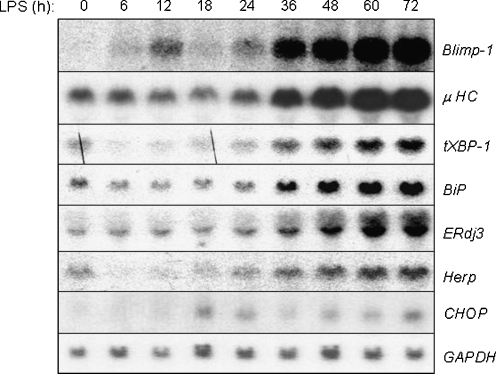
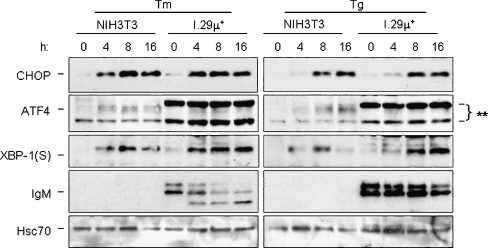
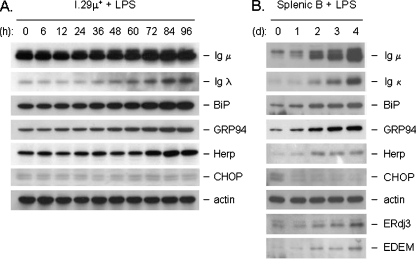
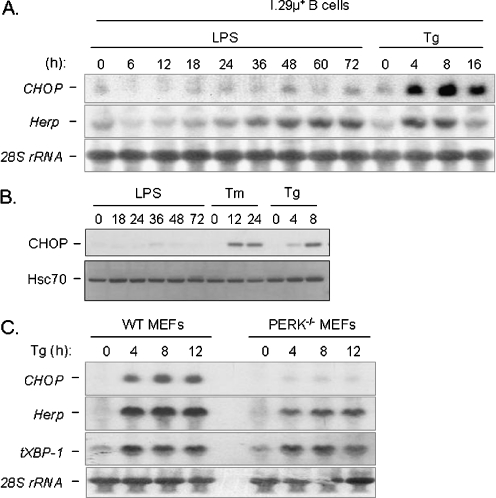
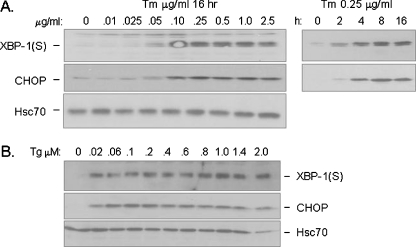
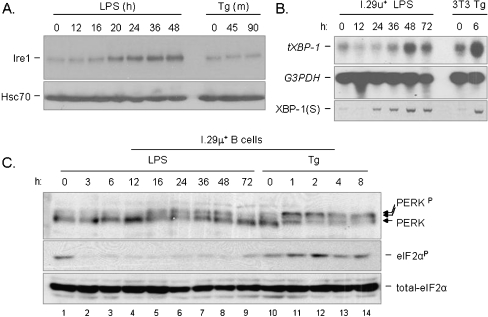
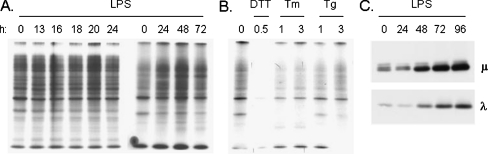
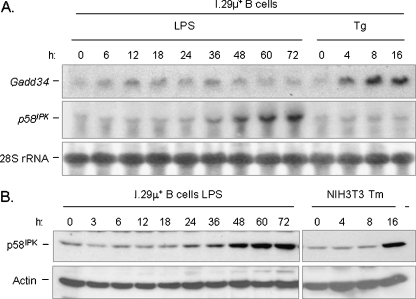
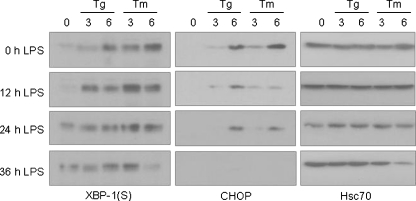
In response to terminal differentiation signals that enable B cells to produce vast quantities of antibodies, a dramatic expansion of the secretory pathway and a corresponding increase in the molecular chaperones and folding enzymes that aid and monitor immunoglobulin synthesis occurs. Recent studies reveal that the unfolded protein response (UPR), which is normally activated by endoplasmic reticulum (ER) stress, plays a critical role in this process. Although B cells activate all three branches of the UPR in response to pharmacological inducers of the pathway, plasma cell differentiation elicits only a partial UPR in which components of the PKR-like ER kinase (PERK) branch are not expressed. This prompted us to further characterize UPR activation during plasma cell differentiation. We found that in response to lipopolysaccharides (LPS)-induced differentiation of the I.29 micro(+) B cell line, Ire1 was activated early, which led to splicing of XBP-1. PERK was partially phosphorylated with similar kinetics, but this was not sufficient to activate its downstream target eIF-2alpha, which initiates translation arrest, or to induce other targets like CHOP or GADD34. Both of these events preceded increased Ig synthesis, arguing this is not the signal for activating these two transducers. Targets of activating transcription factor 6 (ATF6) were up-regulated considerably later, arguing that the ATF6 branch is activated by a distinct signal. Pretreatment with LPS inhibited activation of the PERK branch by pharmacological inducers of the UPR, suggesting that differentiation-induced signals specifically silence this branch. This unique ability to differentially regulate various branches of the UPR allows B cells to accomplish distinct outcomes via the same UPR machinery.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
