

doi: 10.1016/j.jmb.2009.11.003.
Epub 2009 Nov 6.
A novel mechanism of sugar selection utilized by a human X-family DNA polymerase
Affiliations
- PMID: 19900463
- PMCID: PMC2814876
- DOI: 10.1016/j.jmb.2009.11.003
Item in Clipboard
A novel mechanism of sugar selection utilized by a human X-family DNA polymerase
J Mol Biol.
.
Abstract
During DNA synthesis, most DNA polymerases and reverse transcriptases select against ribonucleotides via a steric clash between the ribose 2'-hydroxyl group and the bulky side chain of an active-site residue. In this study, we demonstrated that human DNA polymerase lambda used a novel sugar selection mechanism to discriminate against ribonucleotides, whereby the ribose 2'-hydroxyl group was excluded mostly by a backbone segment and slightly by the side chain of Y505. Such steric clash was further demonstrated to be dependent on the size and orientation of the substituent covalently attached at the ribonucleotide C2'-position.
Copyright 2009 Elsevier Ltd. All rights reserved.
Figures

(A) Sequence alignment of the α-helix M in the thumb domain of each human X-family DNA polymerase. Residue numbers correspond to positions with respect to the N-terminal methionines. The black background indicates which residues are conserved among the family members and the arrow head denotes the potential “steric gate” residue. (B) A close-up view of potential steric interactions with the C2′ position of an incoming ddTTP are depicted in the active site of Polλ (PDB 1XSN). Distances are listed in the table for the Y505 side chain and peptide backbone residues Y505-G508 (purple) near the C2′ position.

(A) Sequence alignment of the α-helix M in the thumb domain of each human X-family DNA polymerase. Residue numbers correspond to positions with respect to the N-terminal methionines. The black background indicates which residues are conserved among the family members and the arrow head denotes the potential “steric gate” residue. (B) A close-up view of potential steric interactions with the C2′ position of an incoming ddTTP are depicted in the active site of Polλ (PDB 1XSN). Distances are listed in the table for the Y505 side chain and peptide backbone residues Y505-G508 (purple) near the C2′ position.

(A) Each D-DNA substrate is composed of a 5′-radiolabeled 21-mer, a 5′-phosphorylated 19-mer, and a 41-mer template where ‘X’ represents A for D-1, C for D-8, G for D-6 and D-6T, and T for D-7. The underlined C:G base pair is A:T for D-6T. (B) A pre-incubated solution of Polλ (120 nM) and 5′-[32P]-labeled D-1 (30 nM) was rapidly mixed with increasing concentrations of UTP·Mg2+ (5 μM, ●; 10 μM, ○; 25 μM, ■; 50 μM, □; 100 μM, ▲; 200 μM, △; 400 μM, ◆ and 800 μM, ◇) for various time intervals. The solid lines are the best fits to a single-exponential equation which determined the observed rate constants, kobs. (C) The kobs values were plotted as a function of UTP concentration. The data (●) were then fit to a hyperbolic equation, yielding a kp of 0.053 ± 0.001 s−1 and a Kd of 114 ± 8 μM.

(A) Each D-DNA substrate is composed of a 5′-radiolabeled 21-mer, a 5′-phosphorylated 19-mer, and a 41-mer template where ‘X’ represents A for D-1, C for D-8, G for D-6 and D-6T, and T for D-7. The underlined C:G base pair is A:T for D-6T. (B) A pre-incubated solution of Polλ (120 nM) and 5′-[32P]-labeled D-1 (30 nM) was rapidly mixed with increasing concentrations of UTP·Mg2+ (5 μM, ●; 10 μM, ○; 25 μM, ■; 50 μM, □; 100 μM, ▲; 200 μM, △; 400 μM, ◆ and 800 μM, ◇) for various time intervals. The solid lines are the best fits to a single-exponential equation which determined the observed rate constants, kobs. (C) The kobs values were plotted as a function of UTP concentration. The data (●) were then fit to a hyperbolic equation, yielding a kp of 0.053 ± 0.001 s−1 and a Kd of 114 ± 8 μM.
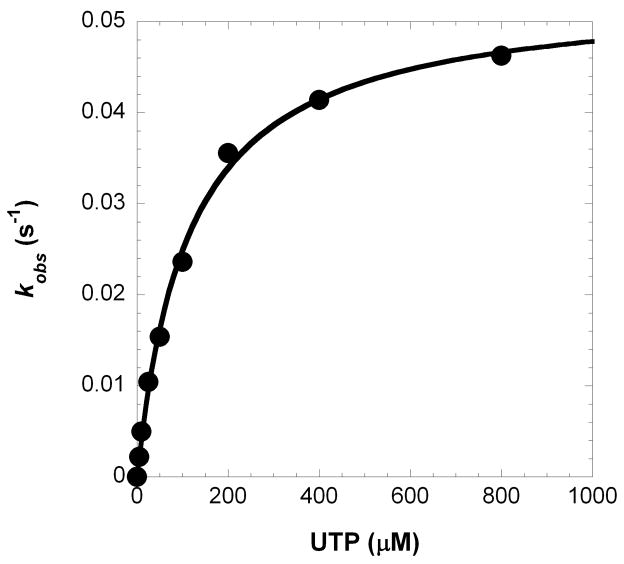
(A) Each D-DNA substrate is composed of a 5′-radiolabeled 21-mer, a 5′-phosphorylated 19-mer, and a 41-mer template where ‘X’ represents A for D-1, C for D-8, G for D-6 and D-6T, and T for D-7. The underlined C:G base pair is A:T for D-6T. (B) A pre-incubated solution of Polλ (120 nM) and 5′-[32P]-labeled D-1 (30 nM) was rapidly mixed with increasing concentrations of UTP·Mg2+ (5 μM, ●; 10 μM, ○; 25 μM, ■; 50 μM, □; 100 μM, ▲; 200 μM, △; 400 μM, ◆ and 800 μM, ◇) for various time intervals. The solid lines are the best fits to a single-exponential equation which determined the observed rate constants, kobs. (C) The kobs values were plotted as a function of UTP concentration. The data (●) were then fit to a hyperbolic equation, yielding a kp of 0.053 ± 0.001 s−1 and a Kd of 114 ± 8 μM.

Using the ternary complex Polλ·gapped DNA·ddTTP (PDB 1XSN), the 2′- and 3′-hydroxyl groups were modeled onto the ribose of the ddTTP substrate. The hydroxyl groups were extracted from the ATP substrate in complex with a reverse transcriptase (PDB 2IAJ). The model presented here showed a steric clash between the 2′-hydroxyl of a dTTP analog (rTTP) and the backbone carboxyl group of Y505.
References
-
- Bonnin A, Lazaro JM, Blanco L, Salas M. A single tyrosine prevents insertion of ribonucleotides in the eukaryotic-type phi29 DNA polymerase. J Mol Biol. 1999;290:241–51. - PubMed
-
- Cases-Gonzalez CE, Gutierrez-Rivas M, Menendez-Arias L. Coupling ribose selection to fidelity of DNA synthesis. The role of Tyr-115 of human immunodeficiency virus type 1 reverse transcriptase. J Biol Chem. 2000;275:19759–67. - PubMed
-
- Yang G, Franklin M, Li J, Lin TC, Konigsberg W. A conserved Tyr residue is required for sugar selectivity in a Pol alpha DNA polymerase. Biochemistry. 2002;41:10256–61. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
