

Key role of T cell defects in age-related vulnerability to West Nile virus
- PMID: 19901080
- PMCID: PMC2806630
- DOI: 10.1084/jem.20090222
Key role of T cell defects in age-related vulnerability to West Nile virus
Abstract
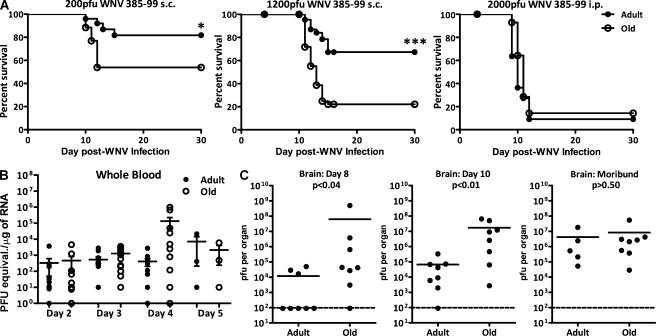
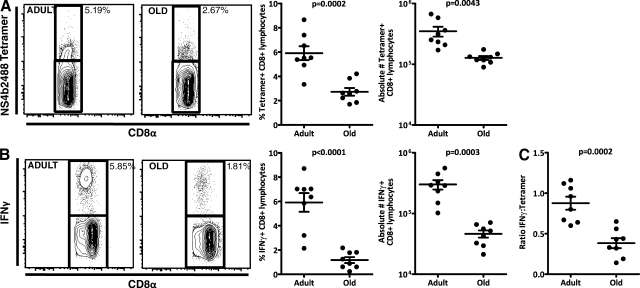
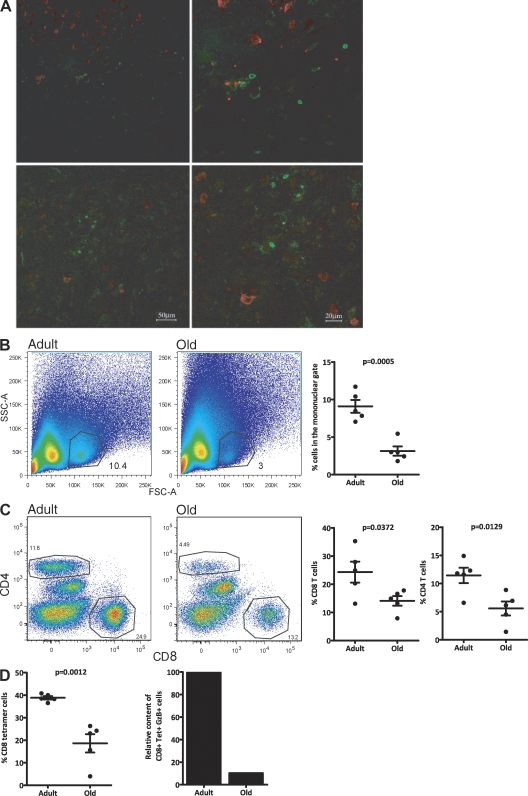
West Nile virus (WNV) infection causes a life-threatening meningoencephalitis that becomes increasingly more prevalent over the age of 50 and is 40-50x more prevalent in people over the age of 70, compared with adults under the age of 40. In a mouse model of age-related vulnerability to WNV, we demonstrate that death correlates with increased viral titers in the brain and that this loss of virus control with age was the result of defects in the CD4 and CD8 T cell response against WNV. Specific age-related defects in T cell responses against dominant WNV epitopes were detected at the level of cytokine and lytic granule production, each of which are essential for resistance against WNV, and in the ability to generate multifunctional anti-WNV effector T cells, which are believed to be critical for robust antiviral immunity. In contrast, at the peak of the response, old and adult T cells exhibited superimposable peptide sensitivity. Most importantly, although the adult CD4 or CD8 T cells readily protected immunodeficient mice upon adoptive transfer, old T cells of either subset were unable to provide WNV-specific protection. Consistent with a profound qualitative and quantitative defect in T cell immunity, old brains contained at least 12x fewer total effector CD8 T cells compared with adult mice at the peak of brain infection. These findings identify potential targets for immunomodulation and treatment to combat lethal WNV infection in the elderly.
Figures



References
-
- Bender B.S., Taylor S.F., Zander D.S., Cottey R. 1995. Pulmonary immune response of young and aged mice after influenza challenge. J. Lab. Clin. Med. 126:169–177 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
