

Efficient operation of NAD(P)H dehydrogenase requires supercomplex formation with photosystem I via minor LHCI in Arabidopsis
- PMID: 19903870
- PMCID: PMC2798312
- DOI: 10.1105/tpc.109.068791
Efficient operation of NAD(P)H dehydrogenase requires supercomplex formation with photosystem I via minor LHCI in Arabidopsis
Abstract
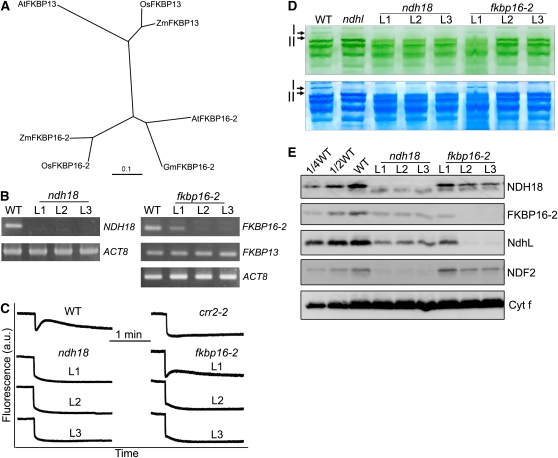
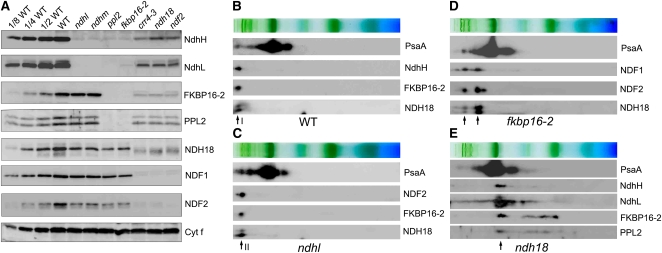
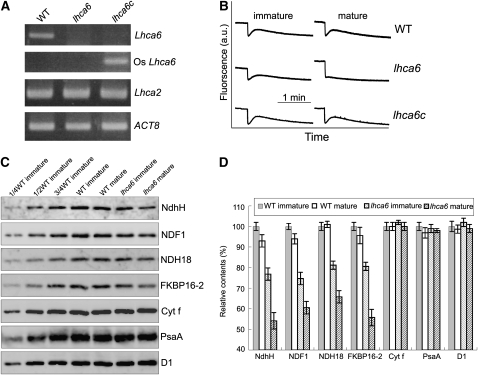
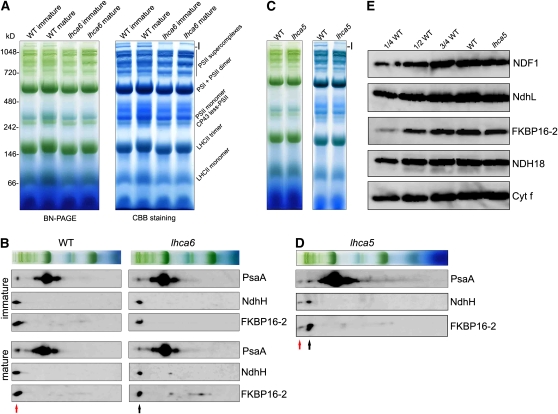
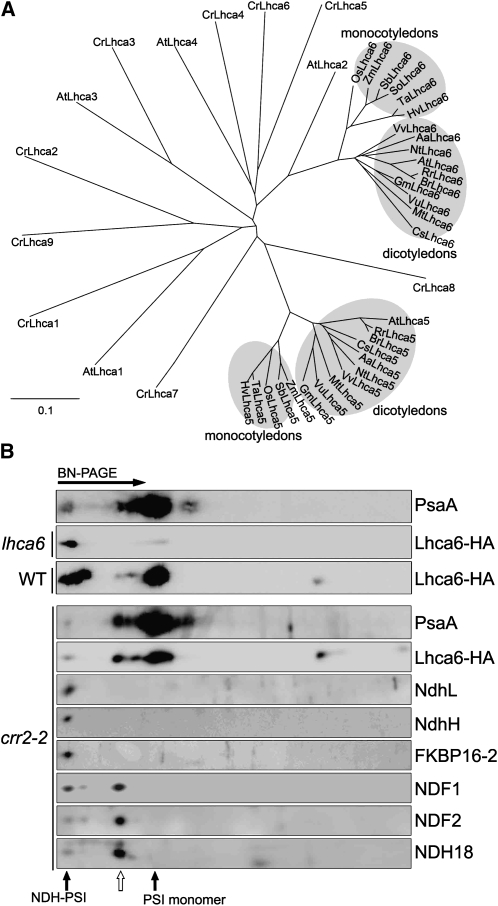
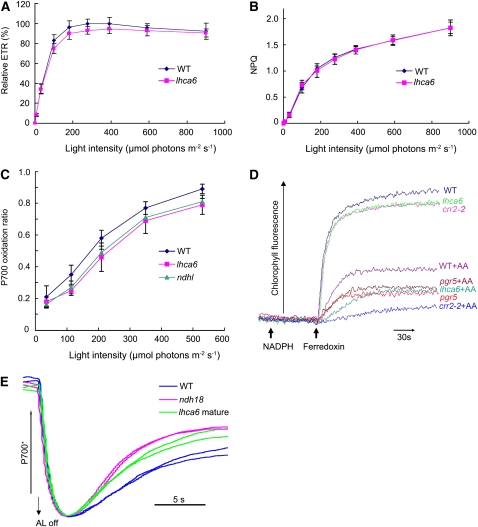
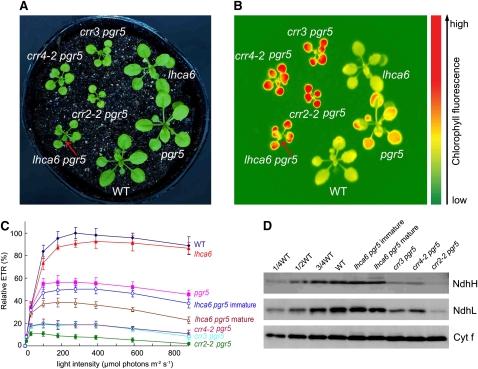
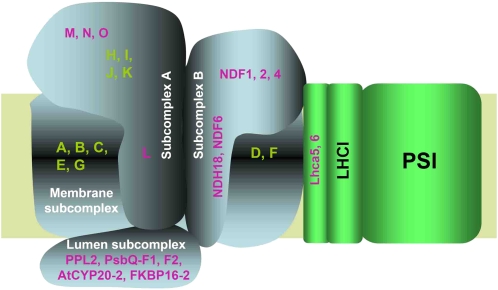
In higher plants, the chloroplast NAD(P)H dehydrogenase (NDH) complex mediates photosystem I (PSI) cyclic and chlororespiratory electron transport. We reported previously that NDH interacts with the PSI complex to form a supercomplex (NDH-PSI). In this study, NDH18 and FKBP16-2 (FK506 Binding Protein 16-2), detected in the NDH-PSI supercomplex by mass spectrometry, were shown to be NDH subunits by the analysis of their knockdown lines. On the basis of extensive mutant characterization, we propose a structural model for chloroplast NDH, whereby NDH is divided into four subcomplexes. The subcomplex A and membrane subcomplex are conserved in cyanobacteria, but the subcomplex B and lumen subcomplex are specific to chloroplasts. Two minor light-harvesting complex I proteins, Lhca5 and Lhca6, were required for the full-size NDH-PSI supercomplex formation. Similar to crr pgr5 double mutants that completely lack cyclic electron flow activity around PSI, the lhca6 pgr5 double mutant exhibited a severe defect in growth. Consistent with the impaired NDH activity, photosynthesis was also severely affected in mature leaves of lhca6 pgr5. We conclude that chloroplast NDH became equipped with the novel subcomplexes and became associated with PSI during the evolution of land plants, and this process may have facilitated the efficient operation of NDH.
Figures

References
-
- Amunts, A., Drory, O., and Nelson, N. (2007). The structure of a plant photosystem I supercomplex at 3.4 A resolution. Nature 447 58–63. - PubMed
-
- Battchikova, N., and Aro, E.-M. (2007). Cyanobacterial NDH-1 complexes: Multiplicity in function and subunit composition. Physiol. Plant. 131 22–32. - PubMed
-
- Battchikova, N., Zhang, P., Rudd, S., Ogawa, T., and Aro, E.-M. (2005). Identification of NdhL and Ssl1690 (NdhO) in NDH-1L and NDH-1M complexes of Synechocystis sp. PCC 6803. J. Biol. Chem. 280 2587–2595. - PubMed
-
- Clough, S.J., and Bent, A.F. (1998). Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16 735–743. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
