

Silver ions increase auxin efflux independently of effects on ethylene response
- PMID: 19903871
- PMCID: PMC2798329
- DOI: 10.1105/tpc.108.065185
Silver ions increase auxin efflux independently of effects on ethylene response
Abstract
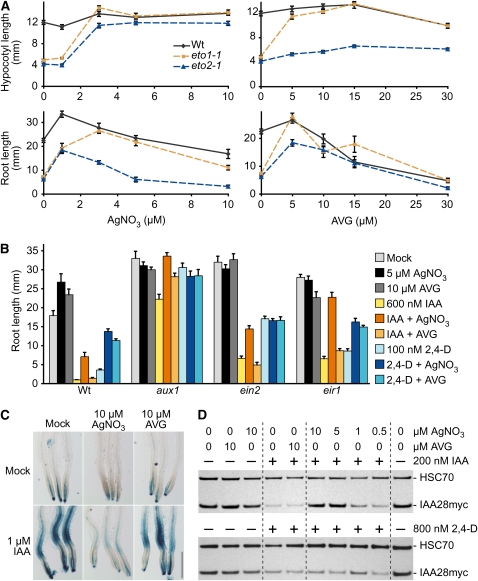
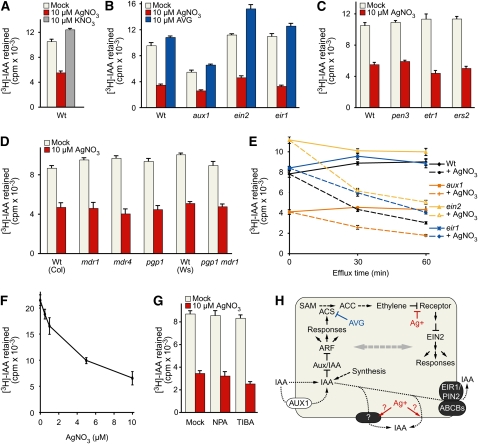
Silver nitrate and aminoethoxyvinylglycine (AVG) are often used to inhibit perception and biosynthesis, respectively, of the phytohormone ethylene. In the course of exploring the genetic basis of the extensive interactions between ethylene and auxin, we compared the effects of silver nitrate (AgNO(3)) and AVG on auxin responsiveness. We found that although AgNO(3) dramatically decreased root indole-3-acetic acid (IAA) responsiveness in inhibition of root elongation, promotion of DR5-beta-glucuronidase activity, and reduction of Aux/IAA protein levels, AVG had more mild effects. Moreover, we found that that silver ions, but not AVG, enhanced IAA efflux similarly in root tips of both the wild type and mutants with blocked ethylene responses, indicating that this enhancement was independent of ethylene signaling. Our results suggest that the promotion of IAA efflux by silver ions is independent of the effects of silver ions on ethylene perception. Although the molecular details of this enhancement remain unknown, our finding that silver ions can promote IAA efflux in addition to blocking ethylene signaling suggest that caution is warranted in interpreting studies using AgNO(3) to block ethylene signaling in roots.
Figures
References
-
- Alonso, J.M., Hirayama, T., Roman, G., Nourizadeh, S., and Ecker, J.R. (1999). EIN2, a bifunctional transducer of ethylene and stress response in Arabidopsis. Science 284 2148–2152. - PubMed
-
- Benavente, L.M., and Alonso, J.M. (2006). Molecular mechanisms of ethylene signaling in Arabidopsis. Mol. Biosyst. 2 165–173. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
