

Memories that last forever: strategies for optimizing vaccine T-cell memory
- PMID: 19903895
- PMCID: PMC2920202
- DOI: 10.1182/blood-2009-06-227546
Memories that last forever: strategies for optimizing vaccine T-cell memory
Abstract
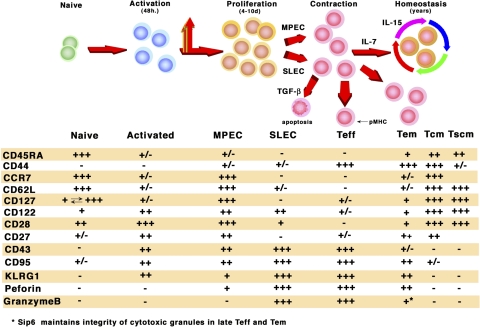
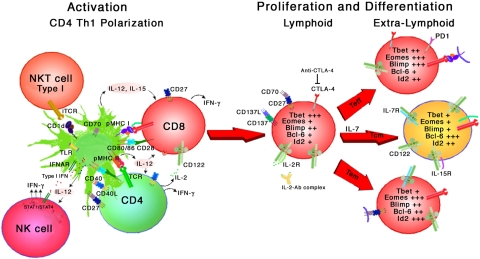
For acute self-limiting infections a vaccine is successful if it elicits memory at least as good as the natural experience; however, for persistent and chronic infections such as HIV, hepatitis C virus (HCV), human papillomavirus (HPV), and human herpes viruses, this paradigm is not applicable. At best, during persistent virus infection the person must be able to maintain the integrity of the immune system in equilibrium with controlling replicating virus. New vaccine strategies are required that elicit both potent high-avidity CD8(+) T-cell effector/memory and central memory responses that can clear the nidus of initial virus-infected cells at mucosal surfaces to prevent mucosal transmission or significantly curtail development of disease. The objective of an HIV-1 T-cell vaccine is to generate functional CD8(+) effector memory cells at mucosal portals of virus entry to prevent viral transmission. In addition, long-lived CD8(+) and CD4(+) central memory cells circulating through secondary lymphoid organs and resident in bone marrow, respectively, are needed to provide a concerted second wave of defense that can contain virus at mucosal surfaces and prevent systemic dissemination. Further understanding of factors which can influence long-lived effector and central memory cell differentiation will significantly contribute to development of effective T-cell vaccines. In this review we will focus on discussing mechanisms involved in T-cell memory and provide promising new approaches toward expanding current vaccine strategies to enhance antiviral memory.
Figures
References
-
- Sallusto F, Lenig D, Förster R, Lipp M, Lanzavecchia A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999;401(6754):708–712. - PubMed
-
- Masopust D, Vezys V, Marzo AL, Lefrancois L. Preferential localization of effector memory cells in nonlymphoid tissue. Science. 2001;291(5512):2413–2417. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
