

Tumor inhibition by sodium selenite is associated with activation of c-Jun NH2-terminal kinase 1 and suppression of beta-catenin signaling
- PMID: 19904745
- PMCID: PMC2862869
- DOI: 10.1002/ijc.25029
Tumor inhibition by sodium selenite is associated with activation of c-Jun NH2-terminal kinase 1 and suppression of beta-catenin signaling
Abstract
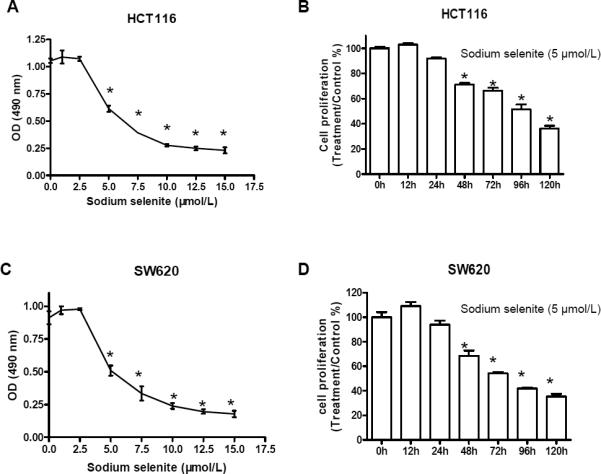
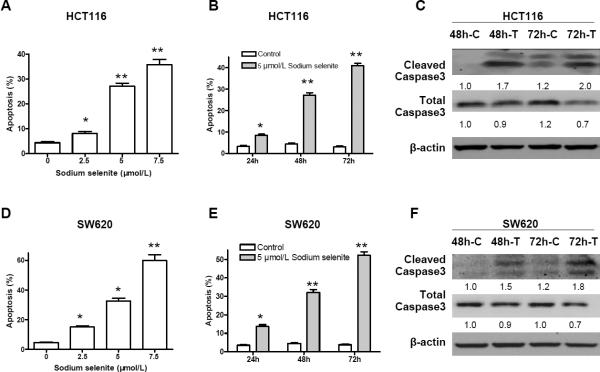
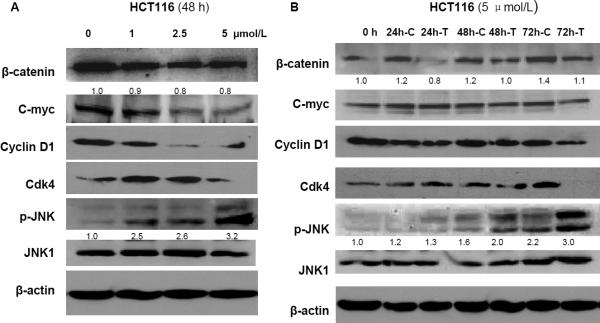
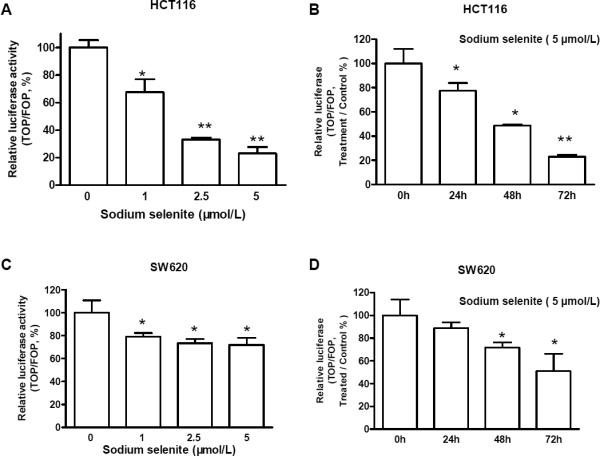
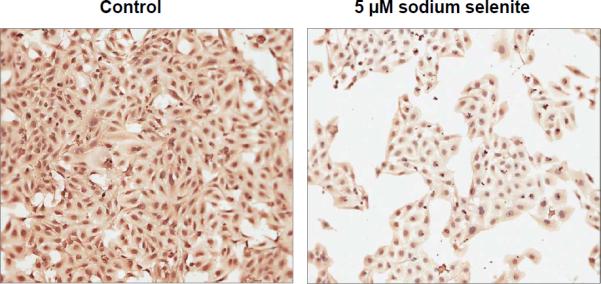
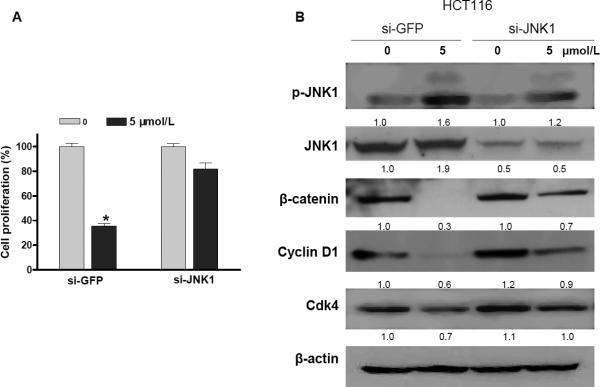
Epidemiological and clinical studies suggest that an increased intake of dietary selenium significantly reduces overall cancer risk, but the anticancer mechanism of selenium is not clear. In this study, we fed intestinal cancer mouse model. Muc2/p21 double mutant mice with a selenium-enriched (sodium selenite) diet for 12 or 24 weeks, and found that sodium selenite significantly inhibited intestinal tumor formation in these animals (p < 0.01), which was associated with phosphorylation of JNK1 and suppression of beta-catenin and COX2. In vitro studies showed that sodium selenite promoted cell apoptosis and inhibited cell proliferation in human colon cancer cell lines HCT116 and SW620. These effects were dose- and time course-dependent, and were also linked to an increase of JNK1 phosphorylation and suppression of beta-catenin signaling. Reduced JNK1 expression by small RNA interference abrogated sufficient activation of JNK1 by sodium selenite, leading to reduced inhibition of the beta-catenin signaling, resulting in reduced efficacy of inhibiting cell proliferation. Taken together, our data demonstrate that sodium selenite inhibits intestinal carcinogenesis in vivo and in vitro through activating JNK1 and suppressing beta-catenin signaling, a novel anticancer mechanism of selenium.
Figures


References
-
- Clark LC, Combs GF, Jr., Turnbull BW, Slate EH, Chalker DK, Chow J, Davis LS, Glover RA, Graham GF, Gross EG, Krongrad A, Lesher JL, Jr., et al. Effects of selenium supplementation for cancer prevention in patients with carcinoma of the skin. A randomized controlled trial. Nutritional Prevention of Cancer Study Group. Jama. 1996;276:1957–63. - PubMed
-
- Greenwald P, Anderson D, Nelson SA, Taylor PR. Clinical trials of vitamin and mineral supplements for cancer prevention. Am J Clin Nutr. 2007;85:314S–7S. - PubMed
-
- Hawk ET, Levin B. Colorectal cancer prevention. J Clin Oncol. 2005;23:378–91. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
