

Dynamic imaging of cannabinoid receptor 1 vesicular trafficking in cultured astrocytes
- PMID: 19906012
- PMCID: PMC2828888
- DOI: 10.1042/AN20090040
Dynamic imaging of cannabinoid receptor 1 vesicular trafficking in cultured astrocytes
Abstract
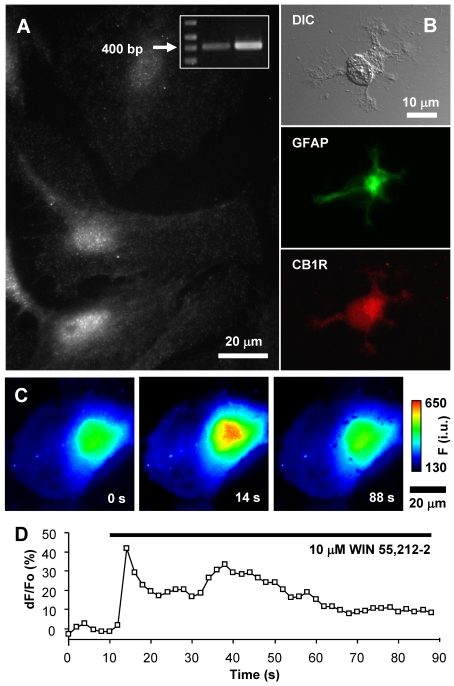
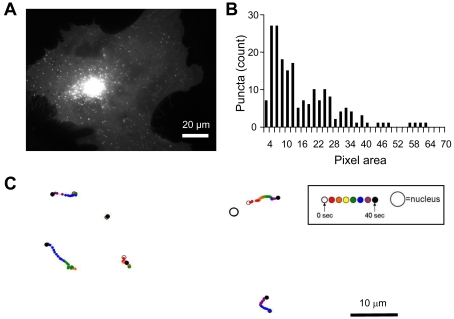
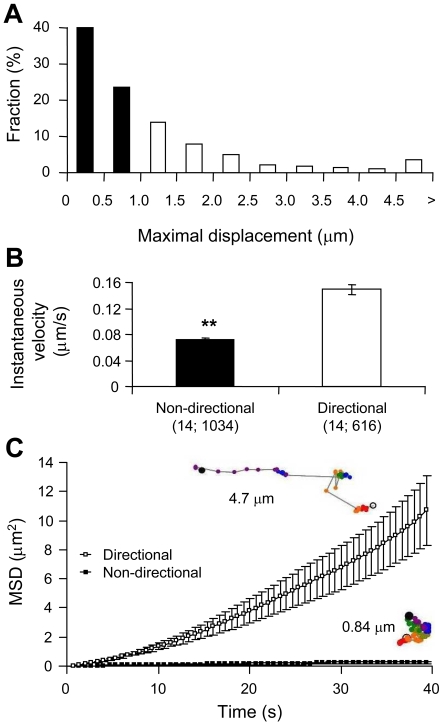
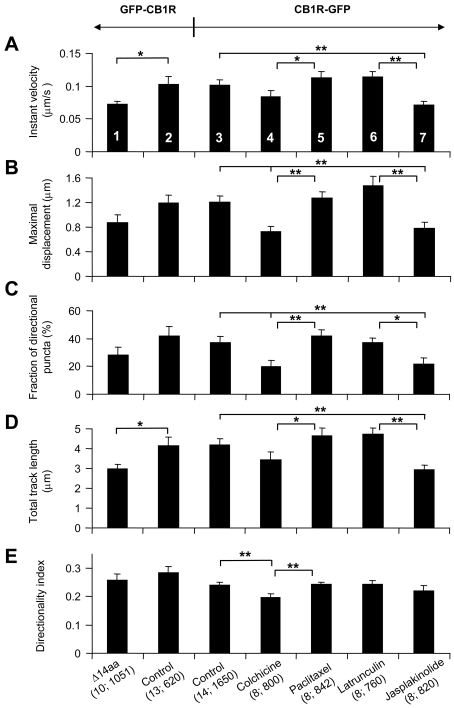
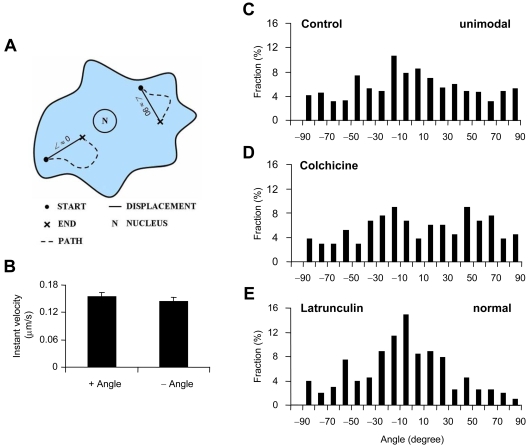
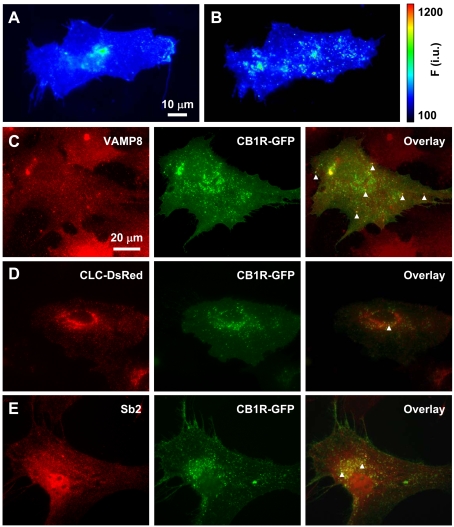
Astrocytes possess GPCRs (G-protein-coupled receptors) for neuroactive substances and can respond via these receptors to signals originating from neurons as well as astrocytes. Like many transmembrane proteins, GPCRs exist in a dynamic equilibrium between receptors expressed at the plasma membrane and those present within intracellular trafficking compartments. The characteristics of GPCR trafficking within astrocytes have not been investigated. We therefore monitored the trafficking of recombinant fluorescent protein chimeras of the CB1R (cannabinoid receptor 1) that is thought to be expressed natively in astrocytes. CB1R chimeras displayed a marked punctate intracellular localization when expressed in cultured rat visual cortex astrocytes, an expression pattern reminiscent of native CB1R expression in these cells. Based upon trafficking characteristics, we found the existence of two populations of vesicular CB1R puncta: (i) relatively immobile puncta with movement characteristic of diffusion and (ii) mobile puncta with movement characteristic of active transport along cytoskeletal elements. The predominant direction of active transport is oriented radially to/from the nuclear region, which can be abolished by disruption of the microtubule cytoskeleton. CB1R puncta are localized within intracellular acidic organelles, mainly co-localizing with endocytic compartments. Constitutive trafficking of CB1R to and from the plasma membrane is an energetically costly endeavour whose function is at present unclear in astrocytes. However, given that intracellular CB1Rs can engage cell signalling pathways, it is likely that this process plays an important regulatory role.
Figures
References
-
- Aoki C, Joh TH, Pickel VM. Ultrastructural localization of β-adrenergic receptor-like immunoreactivity in the cortex and neostriatum of rat brain. Brain Res. 1987;437:264–282. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases