

Dynamic regulation of CD24 and the invasive, CD44posCD24neg phenotype in breast cancer cell lines
- PMID: 19906290
- PMCID: PMC2815544
- DOI: 10.1186/bcr2449
Dynamic regulation of CD24 and the invasive, CD44posCD24neg phenotype in breast cancer cell lines
Abstract
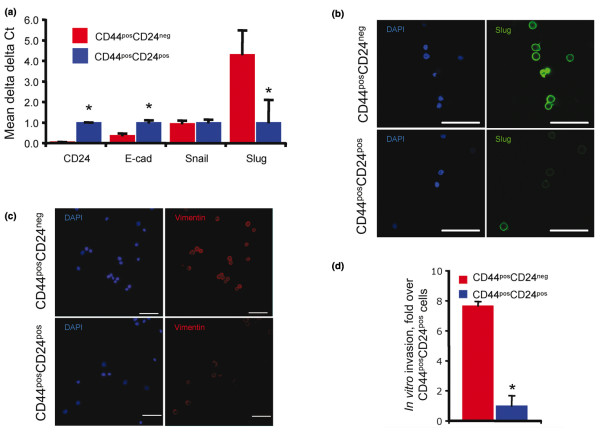
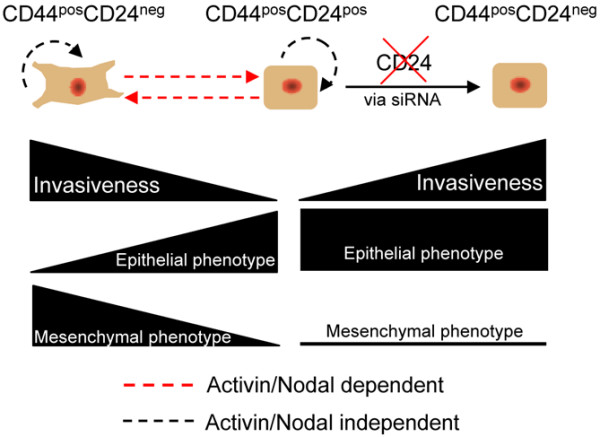
Introduction: The invasive, mesenchymal phenotype of CD44posCD24neg breast cancer cells has made them a promising target for eliminating the metastatic capacity of primary tumors. It has been previously demonstrated that CD44neg/lowCD24pos breast cancer cells lack the ability to give rise to their invasive CD44posCD24neg counterpart. Here we demonstrate that noninvasive, epithelial-like CD44posCD24pos cells readily give rise to invasive, mesenchymal CD44posCD24neg progeny in vivo and in vitro. This interconversion was found to be dependent upon Activin/Nodal signaling.
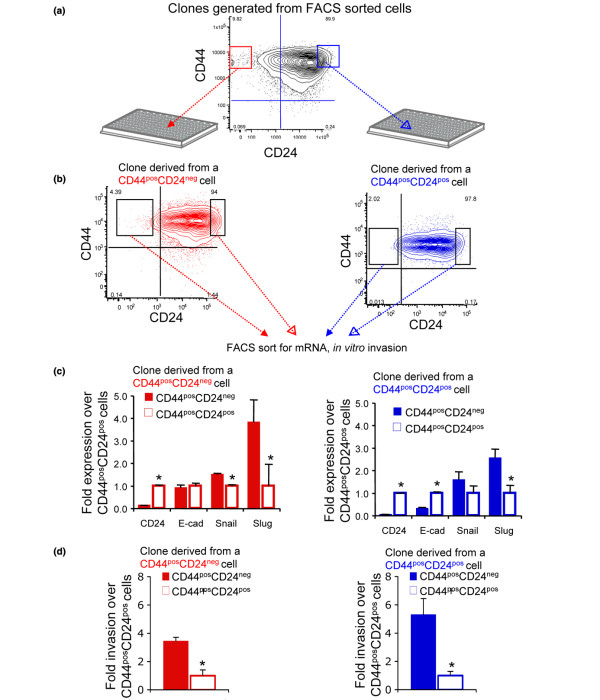
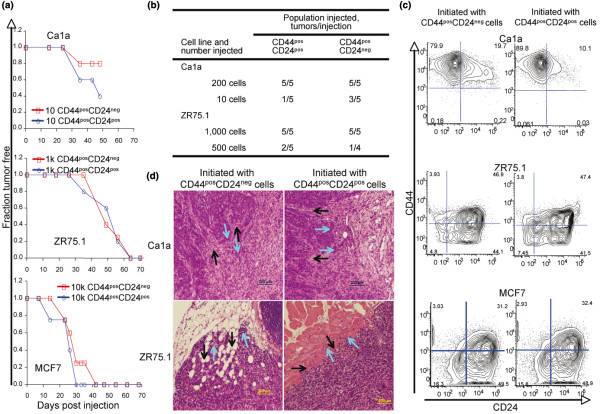
Methods: Breast cancer cell lines were sorted into CD44posCD24pos and CD44posCD24neg populations to evaluate their progeny for the expression of CD44, CD24, and markers of a mesenchymal phenotype. The populations, separated by fluorescence activated cell sorting (FACS) were injected into immunocompromised mice to evaluate their tumorigenicity and invasiveness of the resulting xenografts.
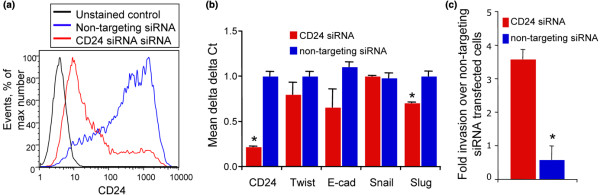
Results: CD24 expression was dynamically regulated in vitro in all evaluated breast cancer cell lines. Furthermore, a single noninvasive, epithelial-like CD44posCD24pos cell had the ability to give rise to invasive, mesenchymal CD44posCD24neg progeny. Importantly, this interconversion occurred in vivo as CD44posCD24pos cells gave rise to xenografts with locally invasive borders as seen in xenografts initiated with CD44posCD24neg cells. Lastly, the ability of CD44posCD24pos cells to give rise to mesenchymal progeny, and vice versa, was blocked upon ablation of Activin/Nodal signaling.
Conclusions: Our data demonstrate that the invasive, mesenchymal CD44posCD24neg phenotype is under dynamic control in breast cancer cell lines both in vitro and in vivo. Furthermore, our observations suggest that therapies targeting CD44posCD24neg tumor cells may have limited success in preventing primary tumor metastasis unless Activin/Nodal signaling is arrested.
Figures


References
-
- Friederichs J, Zeller Y, Hafezi-Moghadam A, Grone HJ, Ley K, Altevogt P. The CD24/P-selectin binding pathway initiates lung arrest of human A125 adenocarcinoma cells. Cancer Res. 2000;60:6714–6722. - PubMed
-
- Fischer GF, Majdic O, Gadd S, Knapp W. Signal transduction in lymphocytic and myeloid cells via CD24, a new member of phosphoinositol-anchored membrane molecules. J Immunol. 1990;144:638–641. - PubMed
-
- Shipitsin M, Campbell LL, Argani P, Weremowicz S, Bloushtain-Qimron N, Yao J, Nikolskaya T, Serebryiskaya T, Beroukhim R, Hu M, Halushka MK, Sukumar S, Parker LM, Anderson KS, Harris LN, Garber JE, Richardson AL, Schnitt SJ, Nikolsky Y, Gelman RS, Polyak K. Molecular definition of breast tumor heterogeneity. Cancer Cell. 2007;11:259–273. doi: 10.1016/j.ccr.2007.01.013. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
