

The shape of motile cells
- PMID: 19906578
- PMCID: PMC2864320
- DOI: 10.1016/j.cub.2009.06.053
The shape of motile cells
Abstract
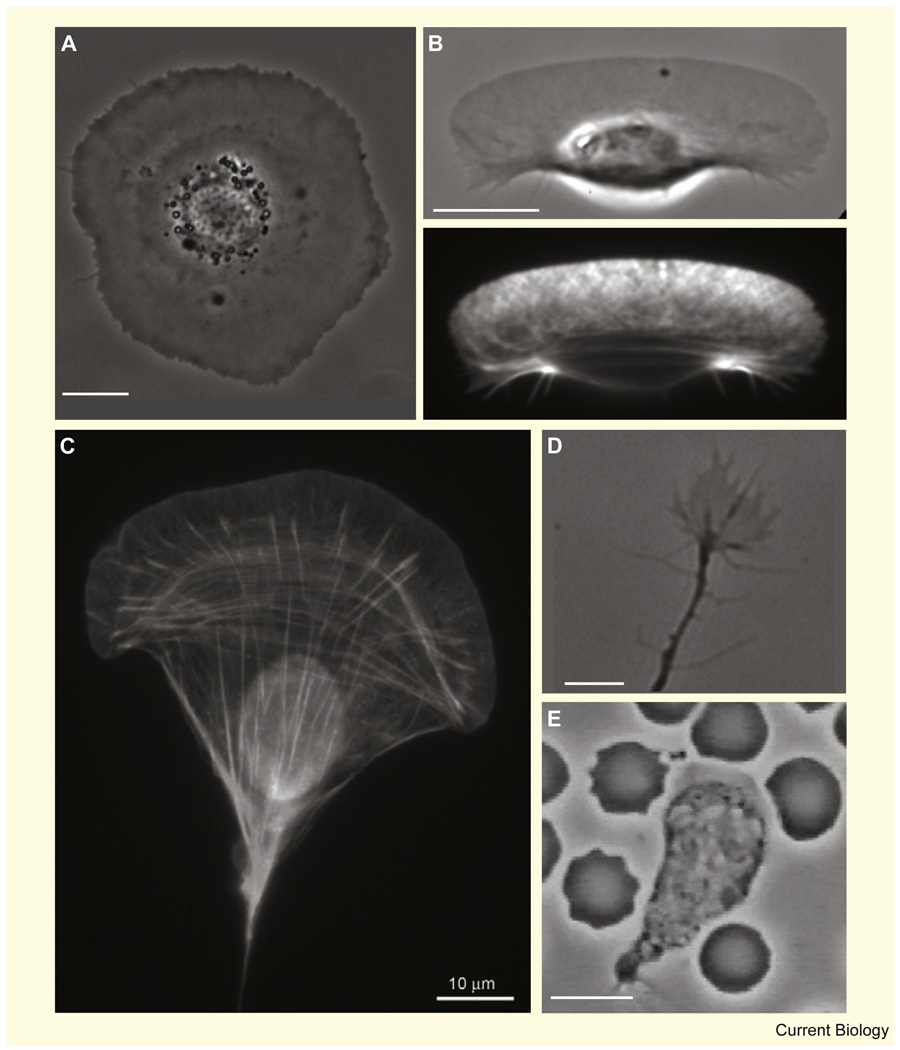
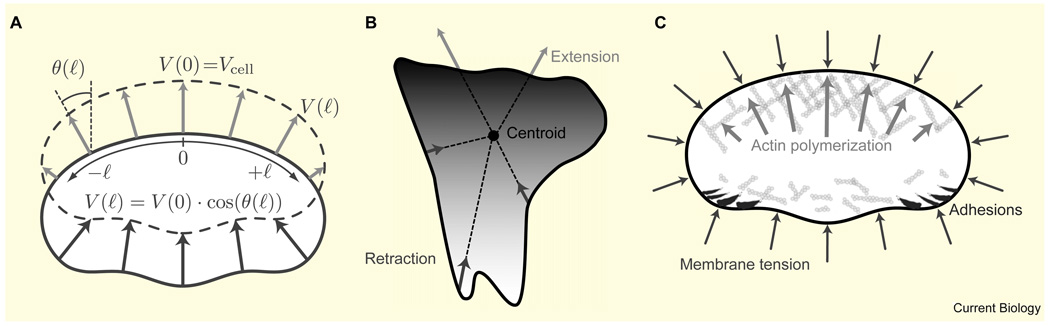
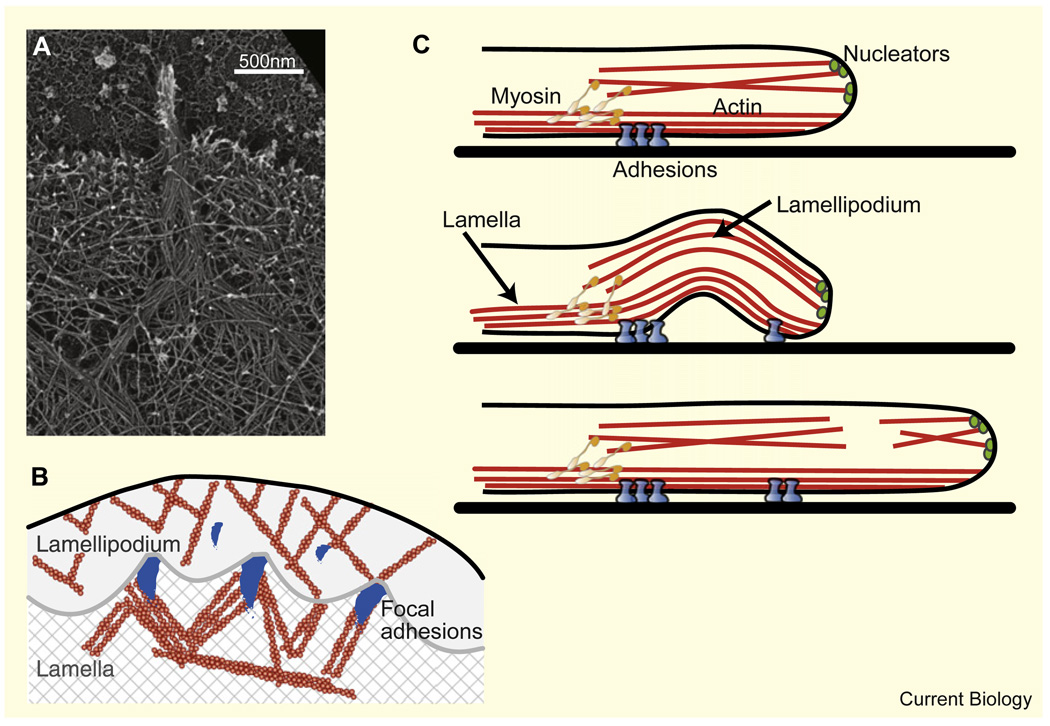
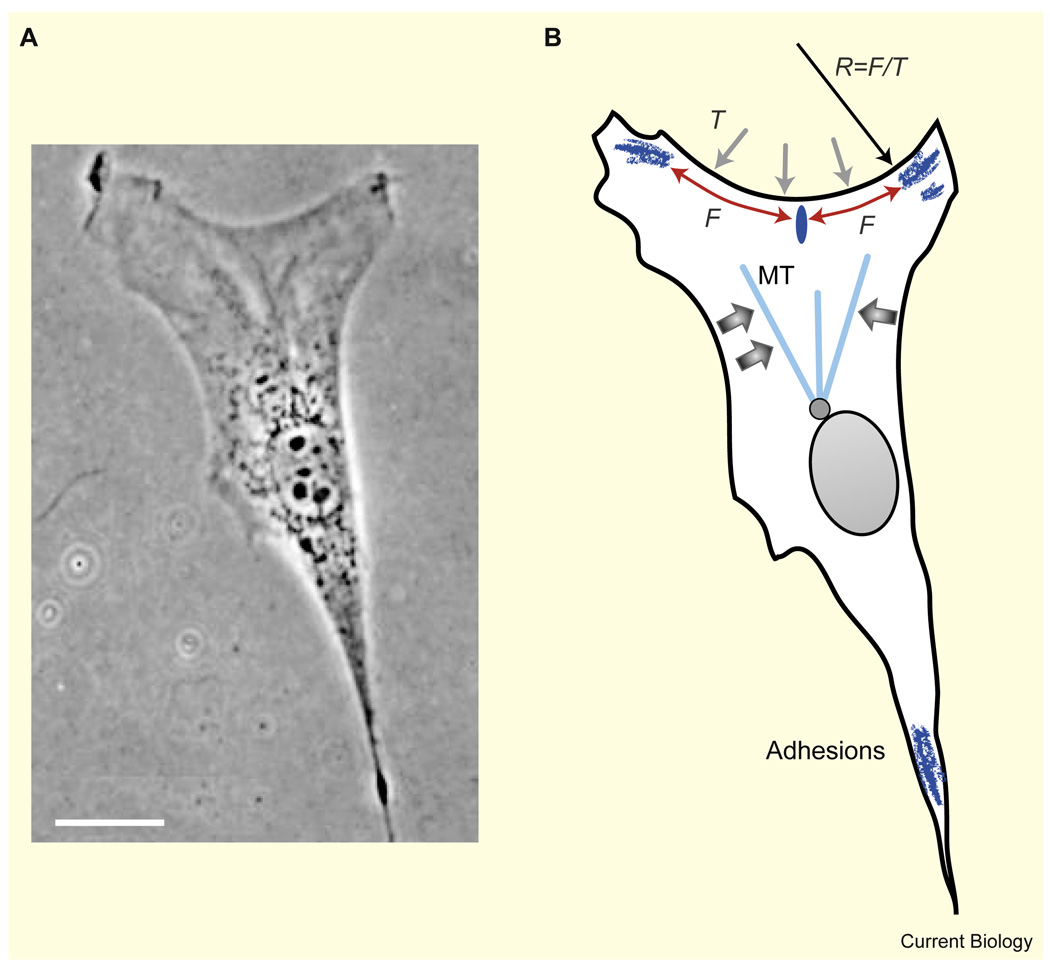
Motile cells - fan-like keratocytes, hand-shaped nerve growth cones, polygonal fibroblasts, to name but a few - come in different shapes and sizes. We discuss the origins of this diversity as well as what shape tells us about the physics and biochemistry underlying cell movement. We start with geometric rules describing cell-edge kinetics that govern cell shape, followed by a discussion of the underlying biophysics; we consider actin treadmilling, actin-myosin contraction, cell-membrane deformations, adhesion, and the complex interactions between these modules, as well as their regulation by microtubules and Rho GTPases. Focusing on several different cell types, including keratocytes and fibroblasts, we discuss how dynamic cell morphology emerges from the interplay between the different motility modules and the environment.
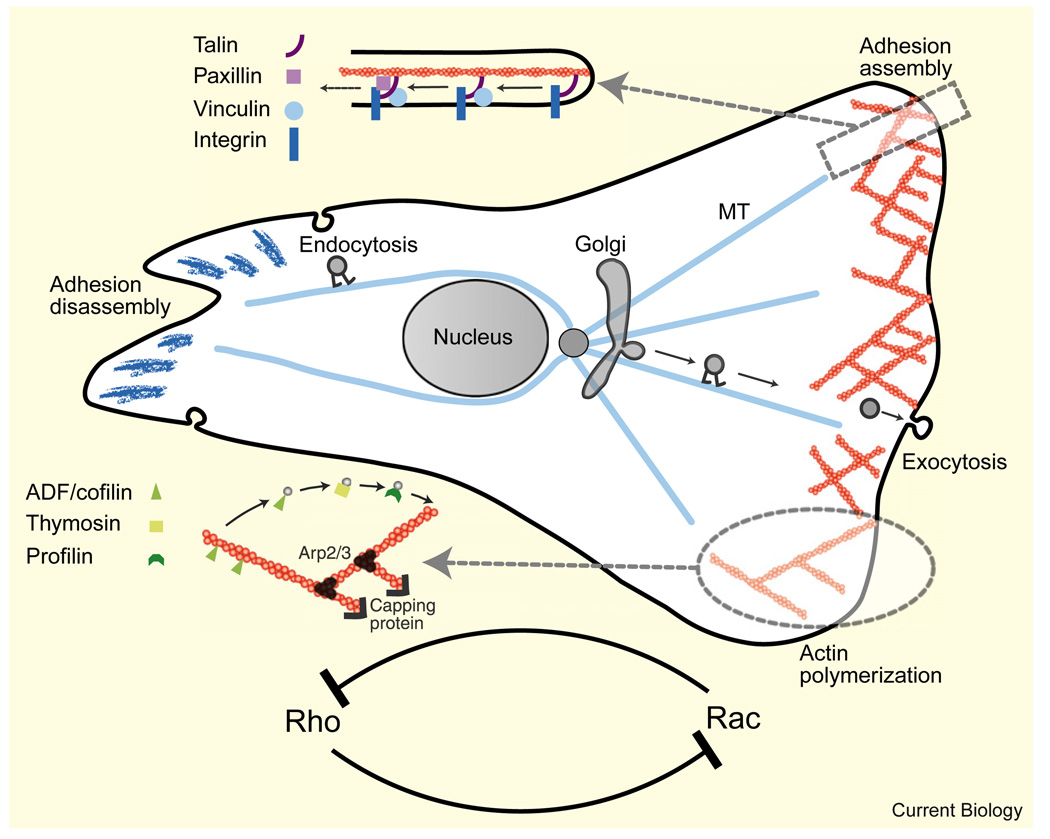
Figures
References
-
- Chen CS, Mrksich M, Huang S, Whitesides GM, Ingber DE. Geometric control of cell life and death. Science. 1997;276:1425–1428. - PubMed
-
- Vasiliev JM. Cytoskeletal mechanisms responsible for invasive migration of neoplastic cells. Int. J. Dev. Biol. 2004;48:425–439. - PubMed
-
- Ridley AJ, Schwartz MA, Burridge K, Firtel RA, Ginsberg MH, Borisy G, Parsons JT, Horwitz AR. Cell migration: integrating signals from front to back. Science. 2003;302:1704–1709. - PubMed
-
- Thery M, Bornens M. Cell shape and cell division. Curr. Opin. Cell Biol. 2006;18:648–657. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
