

Review
doi: 10.1016/j.cub.2009.08.012.
Cell shape and cell division in fission yeast
Affiliations
- PMID: 19906584
- PMCID: PMC2997724
- DOI: 10.1016/j.cub.2009.08.012
Item in Clipboard
Review
Cell shape and cell division in fission yeast
Curr Biol.
.
Abstract
The fission yeast Schizosaccharomyces pombe has served as an important model organism for investigating cellular morphogenesis. This unicellular rod-shaped fission yeast grows by tip extension and divides by medial fission. In particular, microtubules appear to define sites of polarized cell growth by delivering cell polarity factors to the cell tips. Microtubules also position the cell nucleus at the cell middle, marking sites of cell division. Here, we review the microtubule-dependent mechanisms that regulate cell shape and cell division in fission yeast.
Figures
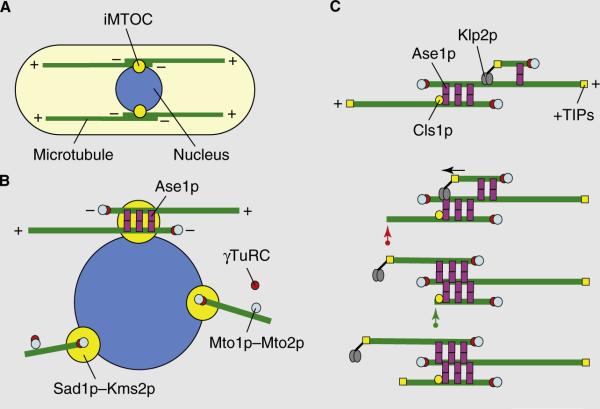
Microtubule organization in fission yeast. (A) A typical fission yeast cell has between three and five dynamic microtubule bundles organized along the long axis of the cell that are organized by iMTOCs into antiparallel bundles with minus ends overlapping at the middle of the cell and plus ends facing and interacting with the cell tips. Two complementary modes of microtubule organization are presented in (B) and (C). (B) In the first model, iMTOCs are tethered to the nuclear membrane. The Mto1p–Mto2p complex, a component of the iMTOC, recruits γ-TuRCs which nucleate microtubules. Microtubule polymers are then bundled into an antiparallel configuration by Ase1p. (C) In the second model, new microtubules nucleate on pre-existing microtubules. The Mto1p–Mto2p complex recruits γ-TuRCs to the lattice of a pre-existing microtubule. Ase1p stabilizes the antiparallel configuration between new and old microtubules. The kinesin Klp2p slides the new microtubule to the minus end of the old microtubule (marked by the arrow), establishing an antiparallel bundle. Microtubule length is regulated by +TIP proteins and the rescue factor Cls1p/Peg1p. A growing microtubule can exhibit catastrophe and shrinkage (red arrow). It can then be rescued by Cls1p/Peg1p at the iMTOC and re-grow (green arrow).
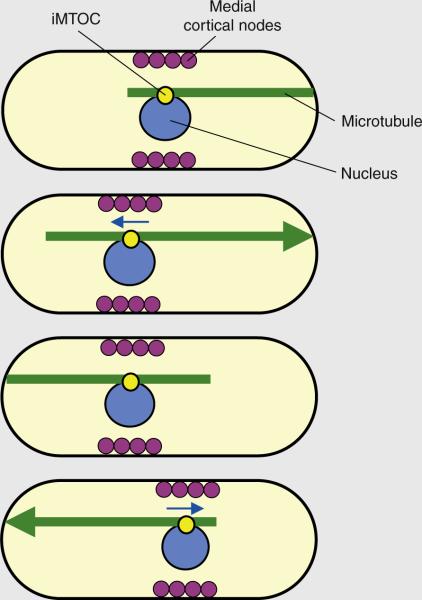
Microtubule pushing centers the nucleus and cortical medial nodes to define the site of cell division. The iMTOC tethers the microtubule bundle to the nucleus. During microtubule–cortex contact, sustained polymerization at the microtubule plus end produces a pushing force (shown by the large arrowhead) that displaces the nucleus in the opposite direction (nuclear movement depicted by blue arrows). The antiparallel configuration of the microtubule bundle ensures that, over time, the nucleus oscillates back and forth toward the geometrical center of a growing cell. Coupling between the nucleus and the medial cortical nodes ensures that the nodes, over time, are also positioned at the cell middle. The medial nodes subsequently organize the actomyosin ring for cell division.
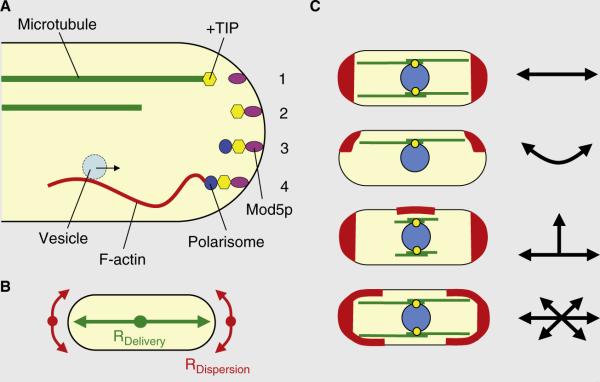
Cell-shape formation in fission yeast. (A) A scheme of microtubule plus ends delivering polarity factors to the cell tip. The microtubule plus end delivers the +TIP complex carrying the Tea1p–Tea4p complex to the tip of the cell (1), where it is docked on the Mod5p receptor (2). The Tea1p–Tea4p complex recruits the polarisome complex containing Bud6p and For3p (3). For3p nucleates F-actin cables, which serve as tracks for the vesicular transport of the growth machinery to the cell tip (4). (B) A model for how the balance between the rate of delivery (RDelivery) and rate of dispersion (RDispersion) of polarity factors defines cell shape. (C) Consequences of changes in the delivery of polarity factors (i.e., RDelivery) or changes in the dispersion of polarity factors (i.e., RDispersion) on cell shape. When RDelivery is equal or greater than RDispersion, cells maintain linear growth. When RDelivery and RDispersion are displaced from the cell long axis, cells grow bent. When RDelivery does not reach the old cell tips, new cell tips are initiated and cells grow T-shaped. Finally, when RDelivery is less than RDispersion, cells grow oval or round.
Similar articles
-
Physical mechanisms redirecting cell polarity and cell shape in fission yeast.Curr Biol. 2008 Nov 25;18(22):1748-53. doi: 10.1016/j.cub.2008.09.047. Curr Biol. 2008. PMID: 19026544 Free PMC article.
-
Fission yeast: in shape to divide.Curr Opin Cell Biol. 2012 Dec;24(6):858-64. doi: 10.1016/j.ceb.2012.10.001. Epub 2012 Nov 3. Curr Opin Cell Biol. 2012. PMID: 23127610 Review.
-
Fission yeast cytoskeletons and cell polarity factors: connecting at the cortex.Biol Cell. 2006 Nov;98(11):619-31. doi: 10.1042/BC20060048. Biol Cell. 2006. PMID: 17042740 Review.
-
Establishing new sites of polarization by microtubules.Curr Biol. 2009 Jan 27;19(2):83-94. doi: 10.1016/j.cub.2008.12.008. Epub 2009 Jan 15. Curr Biol. 2009. PMID: 19147354 Free PMC article.
-
Shaping fission yeast with microtubules.Cold Spring Harb Perspect Biol. 2009 Jul;1(1):a001347. doi: 10.1101/cshperspect.a001347. Cold Spring Harb Perspect Biol. 2009. PMID: 20066076 Free PMC article. Review.
Cited by
-
Processes Controlling the Contractile Ring during Cytokinesis in Fission Yeast, Including the Role of ESCRT Proteins.J Fungi (Basel). 2024 Feb 15;10(2):154. doi: 10.3390/jof10020154. J Fungi (Basel). 2024. PMID: 38392827 Free PMC article. Review.
-
Conserved Orb6 phosphorylation sites are essential for polarized cell growth in Schizosaccharomyces pombe.PLoS One. 2012;7(5):e37221. doi: 10.1371/journal.pone.0037221. Epub 2012 May 21. PLoS One. 2012. PMID: 22629372 Free PMC article.
-
Cells in tight spaces: the role of cell shape in cell function.J Cell Biol. 2010 Oct 18;191(2):233-6. doi: 10.1083/jcb.201009048. J Cell Biol. 2010. PMID: 20956377 Free PMC article.
-
Cytoskeletal dynamics in fission yeast: a review of models for polarization and division.HFSP J. 2010 Jun;4(3-4):122-30. doi: 10.2976/1.3385659. Epub 2010 Apr 15. HFSP J. 2010. PMID: 21119765 Free PMC article.
-
Forces that shape fission yeast cells.Mol Biol Cell. 2017 Jul 7;28(14):1819-1824. doi: 10.1091/mbc.E16-09-0671. Mol Biol Cell. 2017. PMID: 28684607 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
