

Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes
- PMID: 19906667
- PMCID: PMC2842731
- DOI: 10.1098/rspb.2009.1818
Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes
Abstract
Scaling relations among plant traits are both cause and consequence of processes at organ-to-ecosystem scales. The relationship between leaf nitrogen and phosphorus is of particular interest, as both elements are essential for plant metabolism; their limited availabilities often constrain plant growth, and general relations between the two have been documented. Herein, we use a comprehensive dataset of more than 9300 observations of approximately 2500 species from 70 countries to examine the scaling of leaf nitrogen to phosphorus within and across taxonomical groups and biomes. Power law exponents derived from log-log scaling relations were near 2/3 for all observations pooled, for angiosperms and gymnosperms globally, and for angiosperms grouped by biomes, major functional groups, orders or families. The uniform 2/3 scaling of leaf nitrogen to leaf phosphorus exists along a parallel continuum of rising nitrogen, phosphorus, specific leaf area, photosynthesis and growth, as predicted by stoichiometric theory which posits that plants with high growth rates require both high allocation of phosphorus-rich RNA and a high metabolic rate to support the energy demands of macromolecular synthesis. The generality of this finding supports the view that this stoichiometric scaling relationship and the mechanisms that underpin it are foundational components of the living world. Additionally, although abundant variance exists within broad constraints, these results also support the idea that surprisingly simple rules regulate leaf form and function in terrestrial ecosystems.
Figures
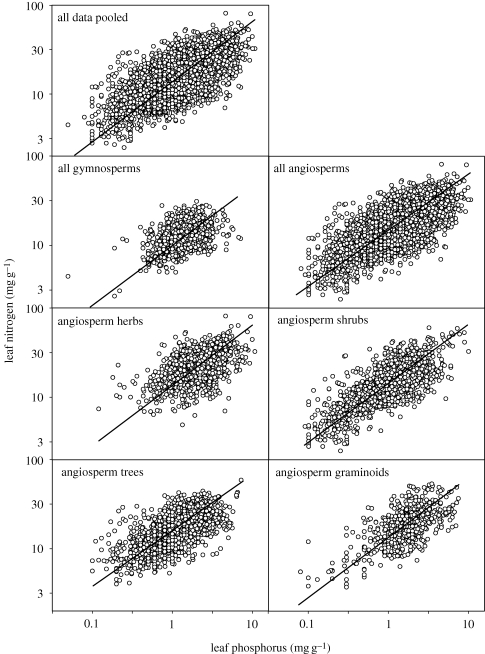
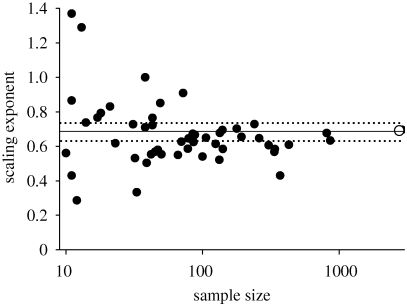
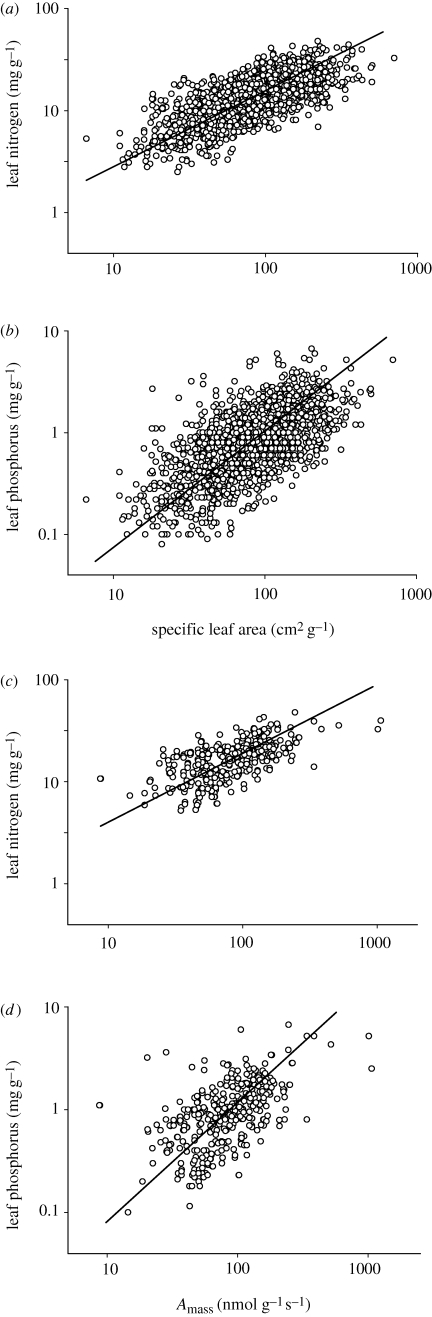
References
-
- Ågren G. I.2004The C:N:P stoichiometry of autotrophs: theory and observations. Ecol. Lett. 7, 185–191 (doi:10.1111/j.1461-0248.2004.00567.x) - DOI
-
- Ågren G. I.2008Stoichiometry and nutrition of plant growth in natural communities. Ann. Rev. Ecol. Evol. Syst. 39, 153–170 (doi:10.1146/annurev.ecolsys.39.110707.173515) - DOI
-
- Chadwick O. A., Derry L. A., Vitousek P. M., Huebert B. J., Hedin L. O.1999Changing sources of nutrients during four million years of ecosystem development. Nature 397, 491–497 (doi:10.1038/17276) - DOI
-
- Elser J. J., et al. 2000Biological stoichiometry from genes to ecosystems. Ecol. Lett. 3, 540–550 (doi:10.1111/j.1461-0248.2000.00185.x) - DOI
-
- Elser J. J., et al. 2003Growth rate—stoichiometry couplings in diverse biota. Ecol. Lett. 6, 936–943 (doi:10.1046/j.1461-0248.2003.00518.x) - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources