

. 2010 Apr 6;7 Suppl 2(Suppl 2):S147-62.
doi: 10.1098/rsif.2009.0411.focus.
Epub 2009 Nov 11.
Cryptochromes--a potential magnetoreceptor: what do we know and what do we want to know?
Affiliations
- PMID: 19906675
- PMCID: PMC2844001
- DOI: 10.1098/rsif.2009.0411.focus
Item in Clipboard
Cryptochromes--a potential magnetoreceptor: what do we know and what do we want to know?
J R Soc Interface.
.
Abstract
Cryptochromes have been suggested to be the primary magnetoreceptor molecules underlying light-dependent magnetic compass detection in migratory birds. Here we review and evaluate (i) what is known about these candidate magnetoreceptor molecules, (ii) what characteristics cryptochrome molecules must fulfil to possibly underlie light-dependent, radical pair based magnetoreception, (iii) what evidence supports the involvement of cryptochromes in magnetoreception, and (iv) what needs to be addressed in future research. The review focuses primarily on our knowledge of cryptochromes in the context of magnetoreception.
Figures
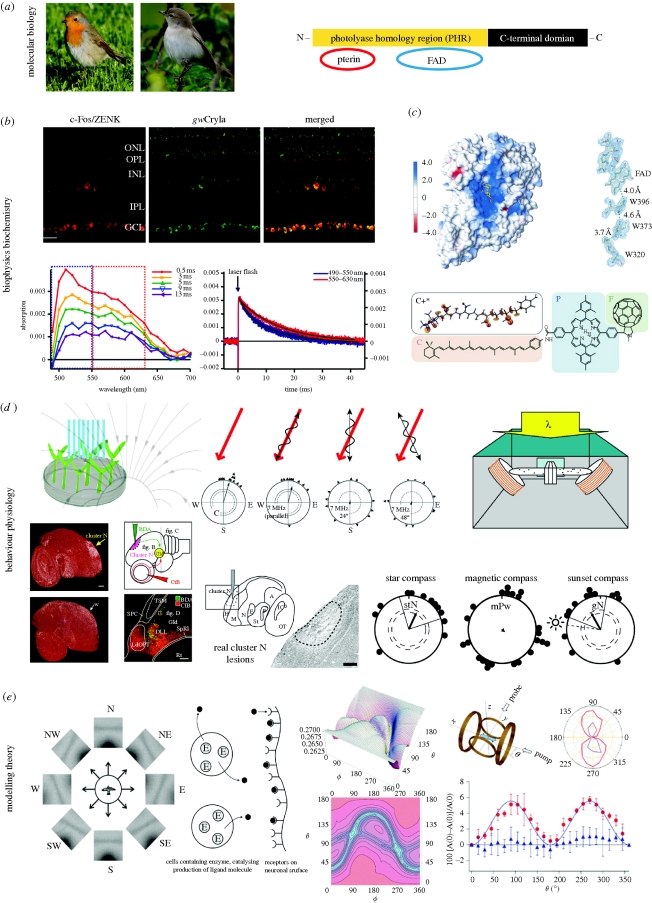
Key evidence suggesting that cryptochromes could function as magnetoreceptors. (a) Photos (©Henrik Mouritsen) show the two migratory model bird species from which, so far, four different members of the cryptochrome multigene family have been detected in the retina (Möller et al. 2004; Mouritsen et al. 2004a; and box 2 of this paper). The schematic illustration indicates the structure of a typical cryptochrome protein (redrawn after Lin & Todo 2005). (b) Cryptochrome has been shown to be expressed in retinal cells that are neuronally active when the birds perform magnetic orientation. The upper panel shows immunohistochemistry staining of a cross section of a garden warbler retina, indicating from left: the expression pattern of neuronal activity marker protiens (c-Fos/ZENK, see main text for details on the use of immediate early genes as neuronal activity markers) in red, the cryptochrome protein expression pattern (which probably included signal from several cryptochromes and not just from gwCry1a as originally thought) in green, and the merged picture which shows colocalization (yellow) of both proteins (reprinted from Mouritsen et al. (2004a). Copyright (2004) National Academy of Sciences, USA). The two graphs show the transient absorption spectra (bottom left) and absorption time profiles (right) of garden warbler cryptochrome 1a. The radical decay rates (blue and red curve, bottom right) indicate that garden warbler cryptochromes form long lived radical species (half-life approx. 10 ms) (reprinted from Liedvogel et al. (2007a)). (c) The first cryptochrome structure presented was cryptochrome-DASH of Synechocystis sp. (reprinted from Brudler et al. (2003). Copyright (2003) with permission from Elsevier (copyright-hyperlink for version published electronically: http://www.cell.com/molecular-cell )); the conserved electron transfer chain is shown at the right. The structure of a migratory bird (or any other animal) cryptochrome remains to be solved. The lower part of (c) shows the structure of the carotenoid–porphyrin–fullerene model system that was designed to demonstrate the feasibility of chemical magnetoreception (reprinted from Maeda et al. (2008). Copyright (2008) with permission from Nature (copyright-hyperlink for version published electronically: http://www.nature.com )). (d) Top left: observations of light-dependent, cryptochrome mediated magnetic field effects on plant growth have been claimed (Ahmad et al. 2007), but unfortunately these findings could not be replicated in a blinded study (Harris et al. 2009; figure redrawn after Ahmad et al. 2007). Top centre: further evidence supporting a radical pair based magnetic compass comes from the observation that weak radiofrequency magnetic fields seem to disrupt the ability of birds to orient by the Earth's magnetic field (Ritz et al. 2004, 2009, 2010; Thalau et al. 2005). The red arrow indicates the orientation of the Earth's magnetic field. The wavy lines indicate the orientation of the oscillating field applied. The circular diagrams show the magnetic orientation of European robins in the magnetic field scenario indicated above the diagram. Each triangle at the circle periphery indicates the mean orientation of an individual bird based on three tests under the given magnetic condition. Arrows indicate the group mean vectors. Inner and outer dashed circles indicate the radii of the group mean vectors needed for directional significance according to the Rayleigh test (inner, p < 0.05; outer, p < 0.01) (redrawn after Ritz et al. (2004). Copyright (2004) with permission from Nature (copyright-hyperlink for version published electronically: http://www.nature.com )). Top right: conditioning experiments on Drosophila fruit flies (setup shown) have recently suggested that a cryptochrome-based magnetosensitive system may also exist in this animal (reprinted from Gegear et al. (2008). Copyright (2008) with permission from Nature (copyright-hyperlink for version published electronically: http://www.nature.com )). Bottom left: in migratory birds, a forebrain area (cluster N) has been identified, which shows light-dependent, movement-independent, neuronal activation at night time when migratory birds performed magnetic orientation under dim light (sagittal brain section through centre of cluster N showing neuronal activation pattern as indicated by ZENK mRNA expression (white signal) at night in a migratory (upper photo) and non-migratory bird (lower photo); Mouritsen et al. 2005; Liedvogel et al. 2007b; Feenders et al. 2008; reprinted from Mouritsen et al. (2005). Copyright (2005) National Academy of Sciences, USA). Bottom centre: a tracing study documented a functional connection between the retinal ganglion cells and cluster N via the thalamofugal visual pathway (schematic drawing). The photo shows that in the visual thalamus, red axons projecting from the eye meet with green backfilled neurons projecting from the thalamus to cluster N (reprinted from Heyers et al. (2007)). Bottom right: the photo shows an example part of a sagitally cut brain section through the centre of cluster N from a cluster N lesioned European robin, stained with a neuroanatomical marker. Notice that the tissue where cluster N should be located is destroyed due to the lesion. The circular diagrams show that cluster N lesioned birds tested in a planetarium simulating the local starry sky (stN, star north) still orient towards the typical north-northeast spring migratory direction, that birds with cluster N lesions could not orient using their magnetic compass (mPw, magnetic poleward), and that birds with cluster N lesions could also orient during sunset, presumably using their sun compass (gN, geographical north). Radial lines indicate the 95% confidence intervals (figure redrawn after Zapka et al. (2009). Copyright (2009) with permission from Nature (copyright-hyperlink for version published electronically: http://www.nature.com )). (e) Theoretical work has shown that, in principle, a radical pair based mechanism could form the basis of magnetic compass orientation (e.g. Ritz et al. 2000; Weaver et al. 2000; Cintolesi et al. 2003; Solov'yov et al. 2007). Left: theoretical visual modulation patterns by the Earth's magnetic field for a bird flying parallel to the horizon looking towards different directions, assuming radical pair receptors oriented perpendicularly to the retina's surface in the bird's eye (reprinted from Ritz et al. (2000). Copyright (2000), with permission from Elsevier (copyright-hyperlink for version published electronically: http://www.cell.com/biophysj )). Centre left: a model for an enzyme (E)-catalysed magnetic field sensitive radical pair reaction. The catalytic reaction produces a ligand (black circles) that leaves the cell and binds to a receptor attached to neural tissue. Subsequent signal transduction depends on the equilibrium between free and unbound ligand (reprinted from Weaver et al. (2000). Copyright (2000) with permission from Nature (copyright-hyperlink for version published electronically: http://www.nature.com )). Centre right: theoretical calculations of radical pair reaction product yields: the picture shows a simulation of the orientation dependence of singlet recombination probability for a flavin–tryptophan radical pair under Earth-strength magnetic fields (B0 = 50 µT, the lifetime of the radical pair is 5µs); orientation dependence is shown both as a three-dimensional representation (top) and as a contour plot (bottom). The direction of the applied magnetic field with respect to the radical pair is defined in terms of the polar angles ϕ and θ (reprinted from Cintolesi et al. (2003). Copyright (2003) with permission from Elsevier (copyright-hyperlink for version published electronically: http://www.elsevier.com/locate/chemphys )). Right: illustration of the chemical compass model (also see c, bottom) providing the proof of principle that a light-induced, radical pair process in a chemical molecule can be sensitive to Earth-strength magnetic fields (Maeda et al. 2008). Right, top left: the experimental setup used to measure the anisotropy of the magnetic field effect: the direction of the applied magnetic field is generated by two orthogonal pairs of Helmholtz coils; the sample alignment along the x-axis is shown by the blue arrow; ϕ is the angle between the magnetic field vector and the x-axis; continuous probe light and pump laser pulses spread along the x and y axes. Top right: a polar plot of the anisotropy of the magnetic field effect as a function of ϕ is plotted for an aligned sample (purple), and (red) by photoselection (i.e. orientation-selective excitation of chromophores by photon absorption). Right, bottom: absorption data from the photoselection measurements (red dots) are plotted as a function of the direction of the applied magnetic field ϕ. As a control, the blue dots confirm that no angular dependence is present in a scenario when the polarization axis of the probe light is vertical (i.e. along z-axis; reprinted from Maeda et al. (2008). Copyright (2008) with permission from Nature (copyright-hyperlink for version published electronically: http://www.nature.com )).

Partial sequence alignment of Cry4 partially coding DNA sequence of the migratory garden warbler Sylvia borin [GQ896539], house sparrow Passer domesticus [AY494987], chicken Gallus gallus [AY102068] and zebra finch Taeniopygia guttata [XM_002198497]. The alignment shows that Cry 4 sequence is highly homologous across species, a common feature of members of the cryptochrome/photolyase family.
References
-
- Adair R. K. 2000. Static and low-frequency magnetic field effects: health risks and therapies. Rep. Prog. Phys. 63, 415–454. (10.1088/0034-4885/63/3/204) - DOI
-
- Ahmad M. 2003. Cryptochromes and flavoprotein blue-light photoreceptors. In Handbook of photochemistry and photobiology (ed. Nalwa H. S.), p. 4 San Diego, CA: Academic Press.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical