

Non-genetic heterogeneity of cells in development: more than just noise
- PMID: 19906852
- PMCID: PMC2778736
- DOI: 10.1242/dev.035139
Non-genetic heterogeneity of cells in development: more than just noise
Abstract
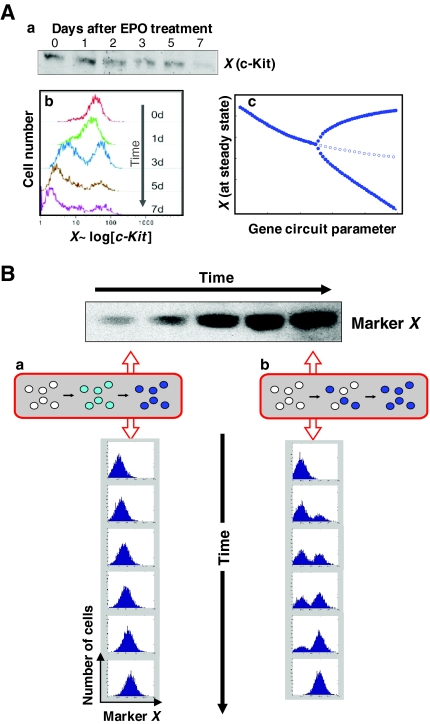
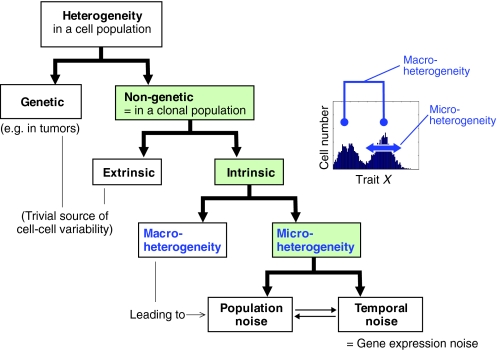
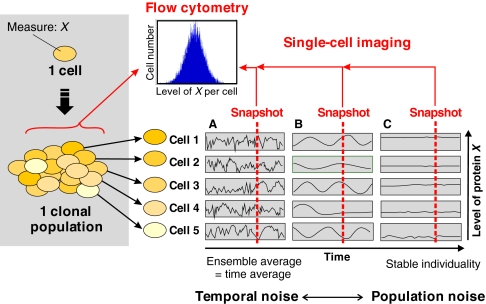
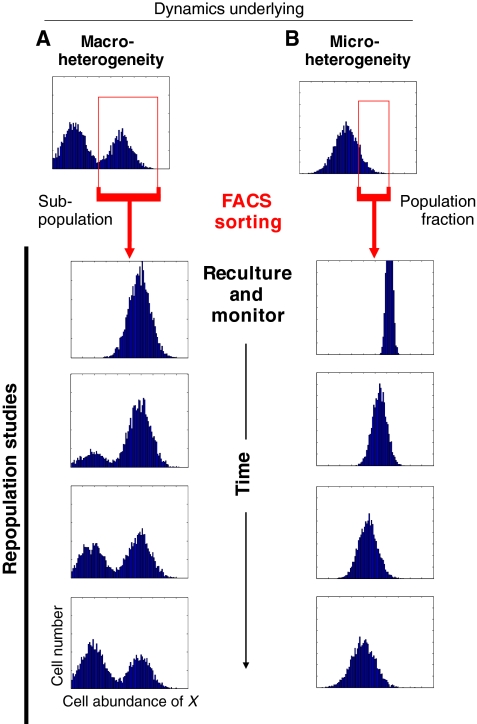
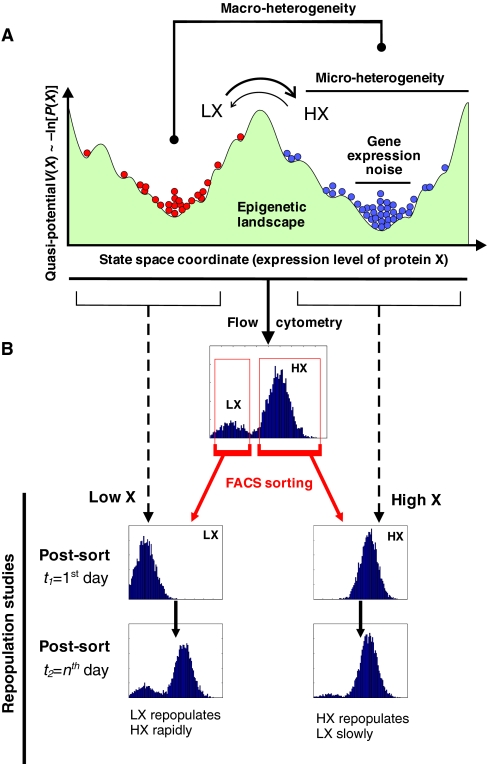
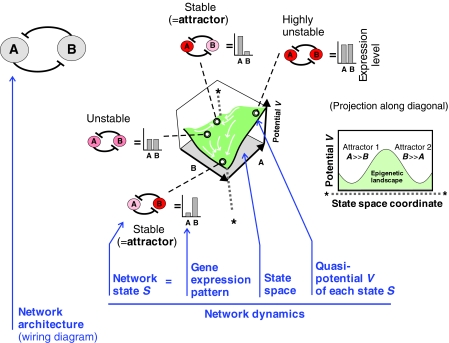
Cell-to-cell variability of gene expression in clonal populations of mammalian cells is ubiquitous. However, because molecular biologists habitually assume uniformity of the cell populations that serve as starting material for experimental analysis, attention to such non-genetic heterogeneity has been scant. As awareness of, and interest in, understanding its biological significance increases, this Primer attempts to clarify the confusing terminologies used in an emerging field that often conflates heterogeneity with noise, and provides a qualitative introduction to the fundamental dynamic principles that underlie heterogeneity. It thus aims to present a useful conceptual framework to organize, analyze and communicate observations made at the resolution of individual cells that indicate that heterogeneity of cell populations plays a biological role, such as in multipotency and cell fate decision.
Figures
References
-
- Aird W. C. (2004). Endothelial cell heterogeneity: a case for nature and nurture. Blood 103, 3994
-
- Aurell E., Sneppen K. (2002). Epigenetics as a first exit problem. Phys. Rev. Lett. 88, 048101 - PubMed
-
- Austin D. W., Allen M. S., McCollum J. M., Dar R. D., Wilgus J. R., Sayler G. S., Samatova N. F., Cox C. D., Simpson M. L. (2006). Gene network shaping of inherent noise spectra. Nature 439, 608-611 - PubMed
-
- Balaban N. Q., Merrin J., Chait R., Kowalik L., Leibler S. (2004). Bacterial persistence as a phenotypic switch. Science 305, 1622-1625 - PubMed
-
- Baldinger K. (1980). Semantic Theory Oxford: Blackwell;
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
