

Review
doi: 10.1242/dev.034876.
Advances in early kidney specification, development and patterning
Affiliations
- PMID: 19906853
- PMCID: PMC2778737
- DOI: 10.1242/dev.034876
Item in Clipboard
Review
Advances in early kidney specification, development and patterning
Development.
2009 Dec.
Abstract
The kidney is a model developmental system for understanding mesodermal patterning and organogenesis, a process that requires regional specification along multiple body axes, the proliferation and differentiation of progenitor cells, and integration with other tissues. Recent progress in the field has highlighted the essential roles of intrinsic nuclear factors and secreted signaling molecules in specifying renal epithelial stem cells and their self-renewal, in driving the complex dynamics of epithelial cell branching morphogenesis, and in nephron patterning. How these developments influence and advance our understanding of kidney development is discussed.
Figures
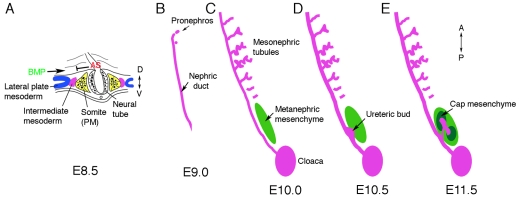
The intermediate mesoderm: its origin and derivatives. In amniotes, the kidney arises from the intermediate mesoderm (IM), between the paraxial somatic mesoderm (PM) and the lateral plate mesoderm (LPM). (A) Schematic cross section through a mouse embryo at embryonic day 8.5 (E8.5) at approximately the sixth somite. The presumptive IM (purple) forms between the LPM (blue) and the PM (yellow). Antagonistic signals (AS) from somites might counteract BMP signals from the LPM to generate the pronephric field. (B-E) Schematics of the IM-derived kidney structures that develop along the anteroposterior (AP) axis in a specific temporal and spatial order. (B) The Wolffian, or pronephric, duct is visible at E9.0 in the mouse and grows caudally by proliferation and extension, inducing epithelial tubules from the adjacent mesenchyme. The pronephros is very rudimentary. (C) Mesonephric tubules at E10, as the nephric duct reaches the cloaca, are more developed in the mid-thoracic region, with a vascularized glomerulus at the proximal end and convoluted tubules draining into the nephric duct. Posterior cells adjacent to the duct form an aggregate called the metanephric mesenchyme (green). (D) By E10.5, an outgrowth of the duct, the ureteric bud (UB), invades the metanephric mesenchyme. (E) By E11.5, the UB has bifurcated and induced mesenchyme (cap mesenchyme) surrounds the tips. Cap mesenchymal cells are the epithelial stem cells of the nephron and generate the glomerular podocyte cells, the parietal epithelium, the proximal tubules, and the distal tubules.
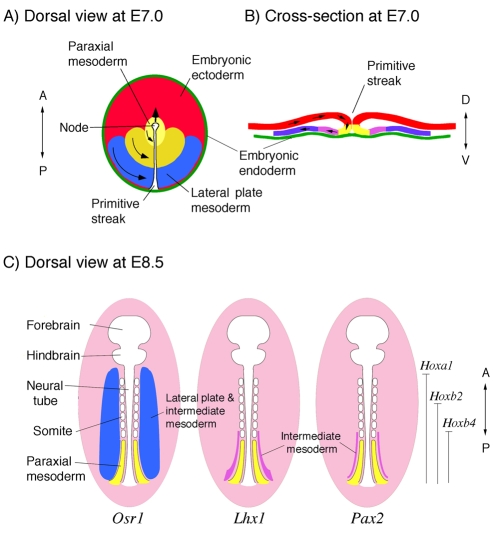
Specification of the intermediate mesoderm. (A,B) During mouse gastrulation, cells of the epiblast ingress through the primitive streak. The more anterior and medial cells are fated to generate paraxial mesoderm (yellow), whereas more posterior and lateral cells make lateral plate mesoderm (blue). The intermediate mesoderm (IM, purple) is not well defined at this time. A is a dorsal view shown as a flat projection, as the mouse epiblast is really cup shaped; B is a posterior cross-section through the primitive streak. (C) Schematic of a flattened E8.5 mouse embryo from the dorsal side. The expression of specific gene products demarcates the IM, which has an anterior border at approximately the sixth somite. Osr1 expression (blue) extends more anterior and more laterally than does Lhx1 expression (purple), which becomes restricted to the IM. The Pax2 (and Pax8) expression domain (purple) also marks the IM and extends caudally. The anterior boundaries of selected Hox genes are marked. The Lhx1 and Pax2 expression domains correspond to the anterior boundary of the Hox4 paralogous group.
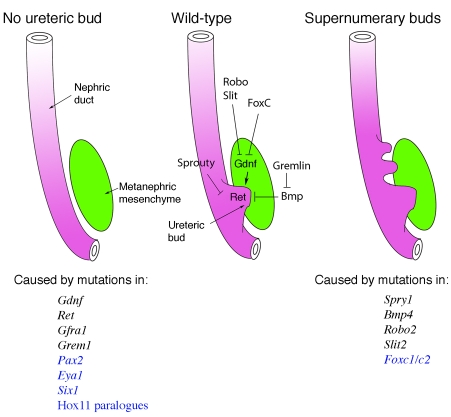
Signals that promote or suppress ureteric bud outgrowth. The outgrowth and invasion of the ureteric bud (UB) from the nephric duct initiates metanephric kidney development. In a wild-type embryo (middle), Gdnf secretion from the metanephric mesenchyme activates the receptor tyrosine kinase Ret, via the co-receptor Gfra1, and promotes UB outgrowth and invasion. Mutations in various regulatory genes can generate two phenotypes, UB ablation (left) or the induction of supernumerary, ectopic UBs (right). Genes encoding signaling proteins are listed in black, those that encode transcription factors are in blue. Not all mutant phenotypes are completely penetrant; for example, Ret and Gdnf mutant mice often show remnants of the UB. Genetic analyses indicate that multiple extracellular inhibitors of Gdnf signaling exist, such as BMPs and Robo/Slit, and an inhibitor of BMPs called gremlin, which is thus an activator of Gdnf signaling. Intracellular inhibitors of Ret signal transduction include Sprouty (Spry1), which may also limit Fgfr signaling. The nuclear factors Foxc1 and Foxc2 act to restrict Gdnf expression to the posterior region, whereas Pax2, Eya1, Hox11 and Six1 proteins all are needed for Gdnf expression.
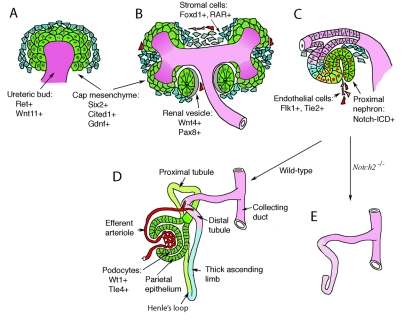
Nephron development and cell lineages. Invasion of the metanephric mesenchyme (green, MM) by the ureteric bud (purple, UB) provides inductive signals that initiate nephrogenesis. (A) UB invasion induces MM cells to condense around the UB tips at E11.5 of mouse development. These so-called cap mesenchymal cells express a unique combination of markers (Six2, Gdnf, Cited1) and define a stem cell population. (B) Cap mesenchyme polarizes into a primitive epithelial sphere, the renal vesicle, coincident with the expression of additional markers, such as Wnt4 and Pax8. Cells in the metanephric mesenchyme that do not aggregate at the bud tips express Foxd1and the retinoic acid receptors (RARs) and mark the stromal population. (C) The renal vesicle fuses to the ureteric stalk, which forms the collecting ducts, and generates an S-shaped body with a proximal and distal cleft. The more proximal cleft is infiltrated by endothelial cells and forms the glomerular tuft. The proximal portion of the S-shaped body activates the Notch pathway, as seen by the presence of the cleaved Notch intracellular domain (ICD). (D) The nephron begins to take shape as glomerular development proceeds and the more proximal tubules elongate and grow towards the medulla to form the descending and ascending limbs of the loop of Henle. (E) Notch signaling is essential for the proximodistal patterning of the nephron, as Notch2 mutations delete all proximal cell types and structures (Cheng et al., 2007).
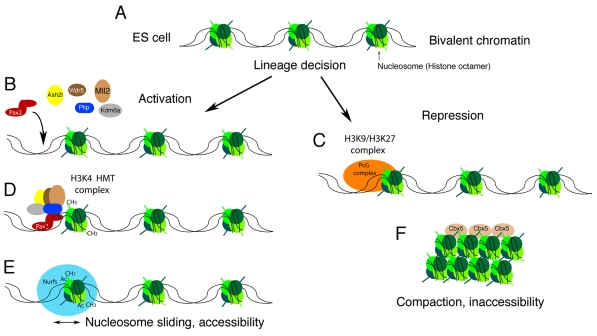
A chromatin model for the epigenetic specification of cell lineages. As cells make lineage decisions, alterations in chromatin structure compartmentalize the genome into active and inactive domains. (A) In pluripotent embryonic stem (ES) cells, tissue-specific genes and developmental regulators are marked with a bivalent histone code that encompasses low levels of both positive and negative histone methylation marks. As cells become specified and their fates are restricted, cell-type specific DNA-binding proteins could provide locus specificity for the modification of chromatin into active (B) or repressed (C) domains. (B) During intermediate mesoderm specification, the Pax2/Pax8 proteins might interact with a histone H3K4 methyltransferase (HMT) complex (D) to prevent repression of kidney-specific genes by the polycomb group (PcG) complexes. (E) High levels of H3K4 trimethylation could then recruit nucleosome remodeling factors (Nurfs) that maintain the accessibility of genes and facilitate transcription. (F) Conversely, PcG-mediated methylation of histone H3 at K9 and K27 could recruit heterochromatin-binding proteins that compact DNA into tightly packaged, silent chromatin. Ac, acetyl; Ash2l, absent small or homeotic like 2; Cbx5, chromobox homolog 5; CH3, methyl; Kdm6a, 4 lysine (K)-specific demethylase 6A; Mll2, mixed-lineage leukemia 2; P, phospho; Ptip, Pax trans-activation domain interacting protein; Wdr5, WD repeat domain 5.
References
-
- Abdelhak S., Kalatzis V., Heilig R., Compain S., Samson D., Vincent C., Weil D., Cruaud C., Sahly I., Leibovici M., et al. (1997). A human homologue of the Drosophila eyes absent gene underlies branchio-oto-renal (BOR) syndrome and identifies a novel gene family. Nat. Genet. 15, 157-164 - PubMed
-
- Azuara V., Perry P., Sauer S., Spivakov M., Jorgensen H. F., John R. M., Gouti M., Casanova M., Warnes G., Merkenschlager M., et al. (2006). Chromatin signatures of pluripotent cell lines. Nat. Cell Biol. 8, 532-538 - PubMed
-
- Barak H., Rosenfelder L., Schultheiss T. M., Reshef R. (2005). Cell fate specification along the anterior-posterior axis of the intermediate mesoderm. Dev. Dyn. 232, 901-914 - PubMed
-
- Basson M. A., Akbulut S., Watson-Johnson J., Simon R., Carroll T. J., Shakya R., Gross I., Martin G. R., Lufkin T., McMahon A. P., et al. (2005). Sprouty1 is a critical regulator of GDNF/RET-mediated kidney induction. Dev. Cell 8, 229-239 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
