

Temporal and spatial dissection of Shh signaling in genital tubercle development
- PMID: 19906863
- PMCID: PMC2778743
- DOI: 10.1242/dev.039768
Temporal and spatial dissection of Shh signaling in genital tubercle development
Abstract
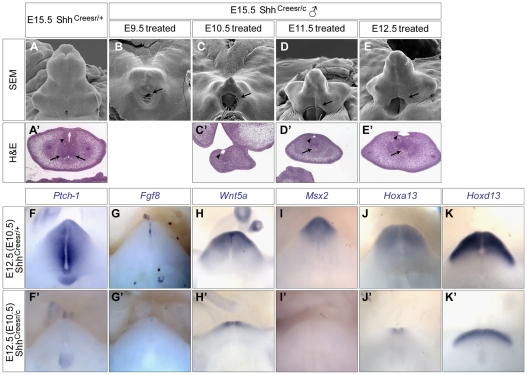
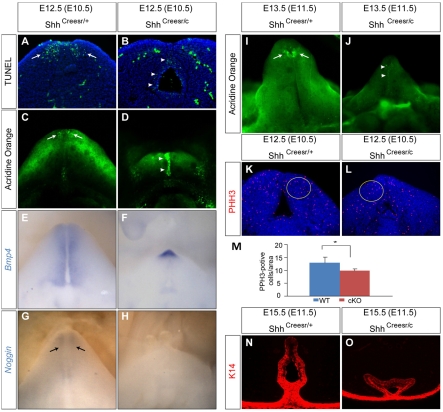
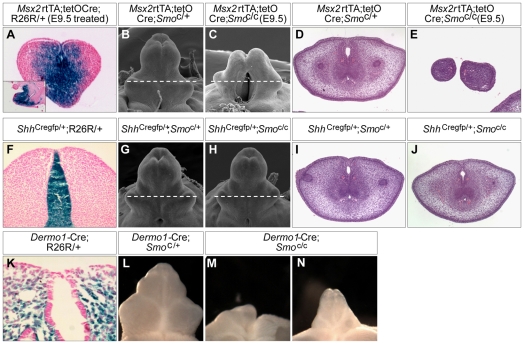
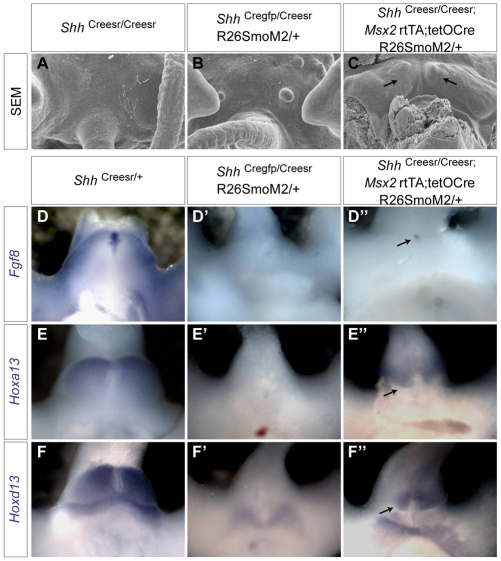
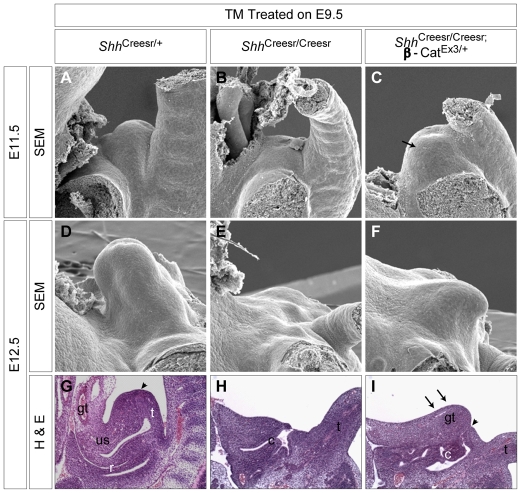
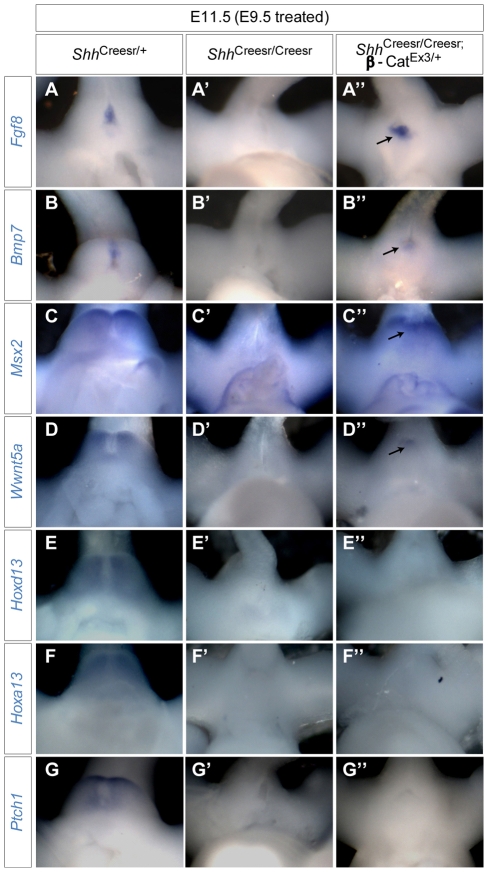
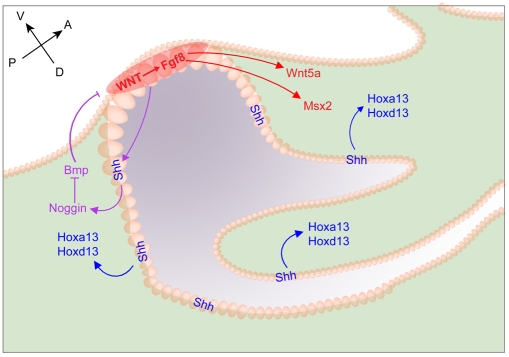
Genital tubercle (GT) initiation and outgrowth involve coordinated morphogenesis of surface ectoderm, cloacal mesoderm and hindgut endoderm. GT development appears to mirror that of the limb. Although Shh is essential for the development of both appendages, its role in GT development is much less clear than in the limb. Here, by removing Shh at different stages during GT development in mice, we demonstrate a continuous requirement for Shh in GT initiation and subsequent androgen-independent GT growth. Moreover, we investigated the Hh responsiveness of different tissue layers by removing or activating its signal transducer Smo with tissue-specific Cre lines, and established GT mesenchyme as the primary target tissue of Shh signaling. Lastly, we showed that Shh is required for the maintenance of the GT signaling center distal urethral epithelium (dUE). By restoring Wnt-Fgf8 signaling in Shh(-/-) cloacal endoderm genetically, we revealed that Shh relays its signal partly through the dUE, but regulates Hoxa13 and Hoxd13 expression independently of dUE signaling. Altogether, we propose that Shh plays a central role in GT development by simultaneously regulating patterning of the cloacal field and supporting an outgrowth signal.
Figures
References
-
- Allard P., Tabin C. J. (2009). Achieving bilateral symmetry during vertebrate limb development. Semin. Cell Dev. Biol. 20, 479-484 - PubMed
-
- Cohn M. J. (2004). Developmental genetics of the external genitalia. Adv. Exp. Med. Biol. 545, 149-157 - PubMed
-
- Echelard Y., Epstein D. J., St-Jacques B., Shen L., Mohler J., McMahon J. A., McMahon A. P. (1993). Sonic hedgehog, a member of a family of putative signaling molecules, is implicated in the regulation of CNS polarity. Cell 75, 1417-1430 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
