

Dosage-dependent hedgehog signals integrated with Wnt/beta-catenin signaling regulate external genitalia formation as an appendicular program
- PMID: 19906864
- PMCID: PMC2778744
- DOI: 10.1242/dev.039438
Dosage-dependent hedgehog signals integrated with Wnt/beta-catenin signaling regulate external genitalia formation as an appendicular program
Abstract
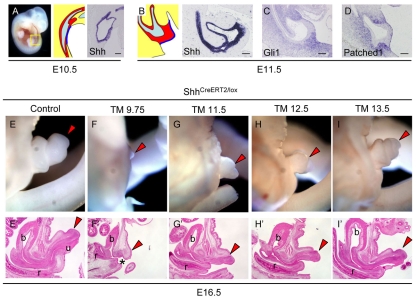
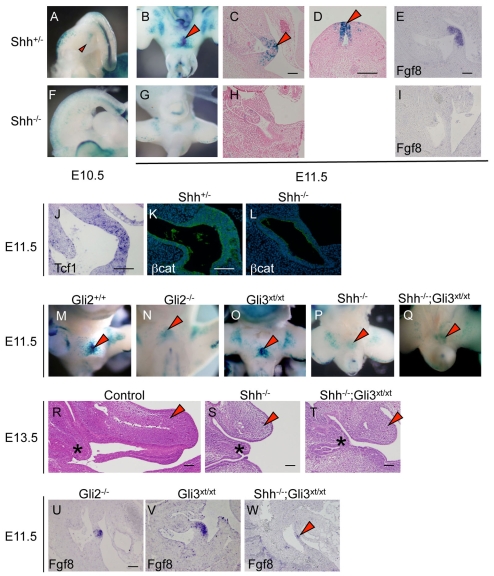
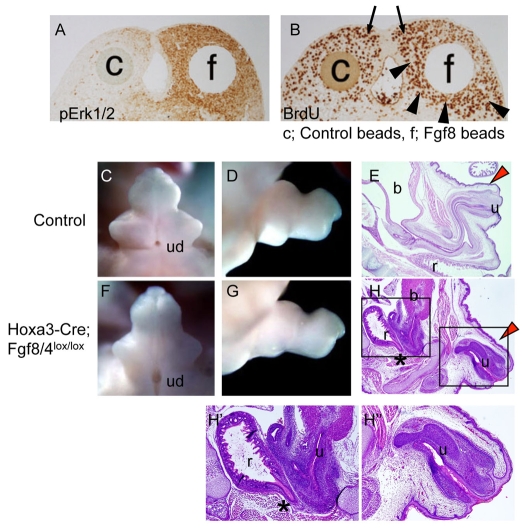
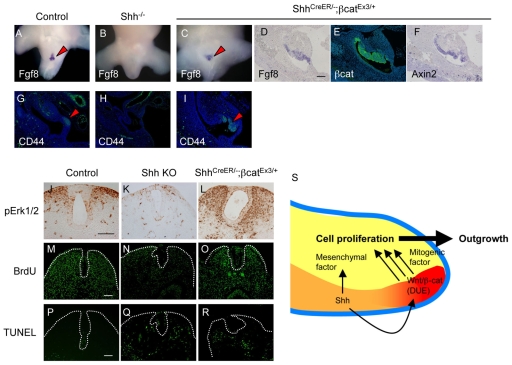
Embryonic appendicular structures, such as the limb buds and the developing external genitalia, are suitable models with which to analyze the reciprocal interactions of growth factors in the regulation of outgrowth. Although several studies have evaluated the individual functions of different growth factors in appendicular growth, the coordinated function and integration of input from multiple signaling cascades is poorly understood. We demonstrate that a novel signaling cascade governs formation of the embryonic external genitalia [genital tubercle (GT)]. We show that the dosage of Shh signal is tightly associated with subsequent levels of Wnt/beta-catenin activity and the extent of external genitalia outgrowth. In Shh-null mouse embryos, both expression of Wnt ligands and Wnt/beta-catenin signaling activity are downregulated. beta-catenin gain-of-function mutation rescues defective GT outgrowth and Fgf8 expression in Shh-null embryos. These data indicate that Wnt/beta-catenin signaling in the distal urethral epithelium acts downstream of Shh signaling during GT outgrowth. The current data also suggest that Wnt/beta-catenin regulates Fgf8 expression via Lef/Tcf binding sites in a 3' conserved enhancer. Fgf8 induces phosphorylation of Erk1/2 and cell proliferation in the GT mesenchyme in vitro, yet Fgf4/8 compound-mutant phenotypes indicate dispensable functions of Fgf4/8 and the possibility of redundancy among multiple Fgfs in GT development. Our results provide new insights into the integration of growth factor signaling in the appendicular developmental programs that regulate external genitalia development.
Figures



References
-
- Ahn K., Mishina Y., Hanks M. C., Behringer R. R., Crenshaw E. B., 3rd (2001). BMPR-IA signaling is required for the formation of the apical ectodermal ridge and dorsal-ventral patterning of the limb. Development 128, 4449-4461 - PubMed
-
- Araki K., Imaizumi T., Okuyama K., Oike Y., Yamamura K. (1997). Efficiency of recombination by Cre transient expression in embryonic stem cells: comparison of various promoters. J. Biochem. 122, 977-982 - PubMed
-
- Beermann F., Kaloulis K., Hofmann D., Murisier F., Bucher P., Trumpp A. (2006). Identification of evolutionarily conserved regulatory elements in the mouse Fgf8 locus. Genesis 44, 1-6 - PubMed
-
- Benazet J. D., Bischofberger M., Tiecke E., Goncalves A., Martin J. F., Zuniga A., Naef F., Zeller R. (2009). A self-regulatory system of interlinked signaling feedback loops controls mouse limb patterning. Science 323, 1050-1053 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
