

Interneuron circuits tune inhibition in retinal bipolar cells
- PMID: 19906884
- PMCID: PMC2807222
- DOI: 10.1152/jn.00458.2009
Interneuron circuits tune inhibition in retinal bipolar cells
Abstract
While connections between inhibitory interneurons are common circuit elements, it has been difficult to define their signal processing roles because of the inability to activate these circuits using natural stimuli. We overcame this limitation by studying connections between inhibitory amacrine cells in the retina. These interneurons form spatially extensive inhibitory networks that shape signaling between bipolar cell relay neurons to ganglion cell output neurons. We investigated how amacrine cell networks modulate these retinal signals by selectively activating the networks with spatially defined light stimuli. The roles of amacrine cell networks were assessed by recording their inhibitory synaptic outputs in bipolar cells that suppress bipolar cell output to ganglion cells. When the amacrine cell network was activated by large light stimuli, the inhibitory connections between amacrine cells unexpectedly depressed bipolar cell inhibition. Bipolar cell inhibition elicited by smaller light stimuli or electrically activated feedback inhibition was not suppressed because these stimuli did not activate the connections between amacrine cells. Thus the activation of amacrine cell circuits with large light stimuli can shape the spatial sensitivity of the retina by limiting the spatial extent of bipolar cell inhibition. Because inner retinal inhibition contributes to ganglion cell surround inhibition, in part, by controlling input from bipolar cells, these connections may refine the spatial properties of the retinal output. This functional role of interneuron connections may be repeated throughout the CNS.
Figures
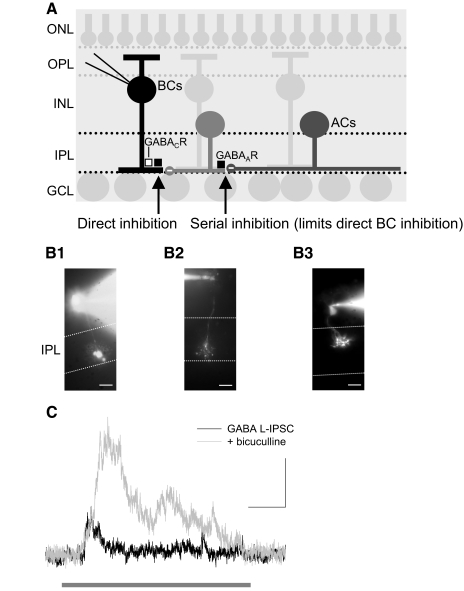
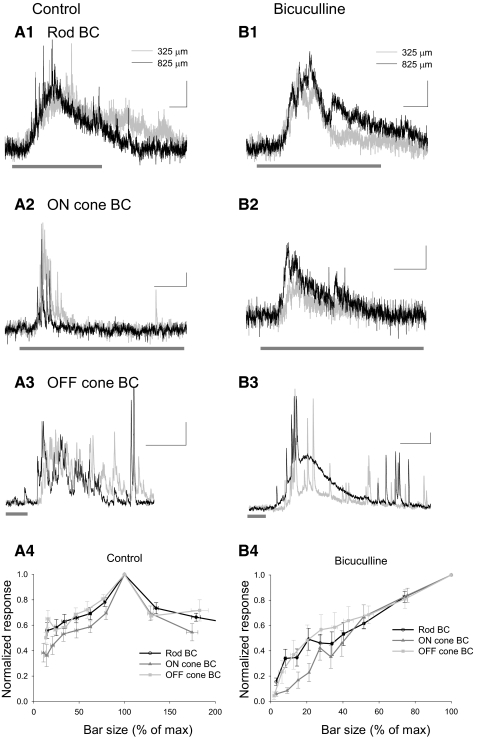
 ). Scale bars are 200 ms and 5 pA.
). Scale bars are 200 ms and 5 pA.





Similar articles
-
Inhibition to retinal rod bipolar cells is regulated by light levels.J Neurophysiol. 2013 Jul;110(1):153-61. doi: 10.1152/jn.00872.2012. Epub 2013 Apr 17. J Neurophysiol. 2013. PMID: 23596335 Free PMC article.
-
GABA(A), GABA(C) and glycine receptor-mediated inhibition differentially affects light-evoked signalling from mouse retinal rod bipolar cells.J Physiol. 2006 Apr 1;572(Pt 1):215-25. doi: 10.1113/jphysiol.2005.103648. Epub 2006 Jan 26. J Physiol. 2006. PMID: 16439422 Free PMC article.
-
Presynaptic inhibition differentially shapes transmission in distinct circuits in the mouse retina.J Physiol. 2007 Jul 15;582(Pt 2):569-82. doi: 10.1113/jphysiol.2007.131763. Epub 2007 Apr 26. J Physiol. 2007. PMID: 17463042 Free PMC article.
-
Amacrine cell-mediated input to bipolar cells: variations on a common mechanistic theme.Vis Neurosci. 2012 Jan;29(1):41-9. doi: 10.1017/S0952523811000241. Vis Neurosci. 2012. PMID: 22310371 Review.
-
Multiple pathways of inhibition shape bipolar cell responses in the retina.Vis Neurosci. 2011 Jan;28(1):95-108. doi: 10.1017/S0952523810000209. Epub 2010 Oct 8. Vis Neurosci. 2011. PMID: 20932357 Free PMC article. Review.
Cited by
-
Glycine transporter 1 modulates GABA release from amacrine cells by controlling occupancy of coagonist binding site of NMDA receptors.J Neurophysiol. 2013 Sep;110(6):1393-403. doi: 10.1152/jn.00193.2013. Epub 2013 Jun 26. J Neurophysiol. 2013. PMID: 23803324 Free PMC article.
-
Inhibition to retinal rod bipolar cells is regulated by light levels.J Neurophysiol. 2013 Jul;110(1):153-61. doi: 10.1152/jn.00872.2012. Epub 2013 Apr 17. J Neurophysiol. 2013. PMID: 23596335 Free PMC article.
-
ON cone bipolar cell axonal synapses in the OFF inner plexiform layer of the rabbit retina.J Comp Neurol. 2013 Apr 1;521(5):977-1000. doi: 10.1002/cne.23244. J Comp Neurol. 2013. PMID: 23042441 Free PMC article.
-
Dopamine D1 receptor modulation of calcium channel currents in horizontal cells of mouse retina.J Neurophysiol. 2016 Aug 1;116(2):686-97. doi: 10.1152/jn.00990.2015. Epub 2016 May 18. J Neurophysiol. 2016. PMID: 27193322 Free PMC article.
-
Independent control of reciprocal and lateral inhibition at the axon terminal of retinal bipolar cells.J Physiol. 2013 Aug 15;591(16):3833-51. doi: 10.1113/jphysiol.2013.253179. Epub 2013 May 20. J Physiol. 2013. PMID: 23690563 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources