

NS1' of flaviviruses in the Japanese encephalitis virus serogroup is a product of ribosomal frameshifting and plays a role in viral neuroinvasiveness
- PMID: 19906906
- PMCID: PMC2812330
- DOI: 10.1128/JVI.01979-09
NS1' of flaviviruses in the Japanese encephalitis virus serogroup is a product of ribosomal frameshifting and plays a role in viral neuroinvasiveness
Abstract
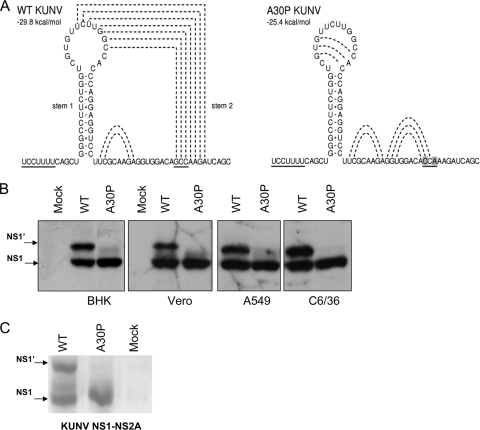
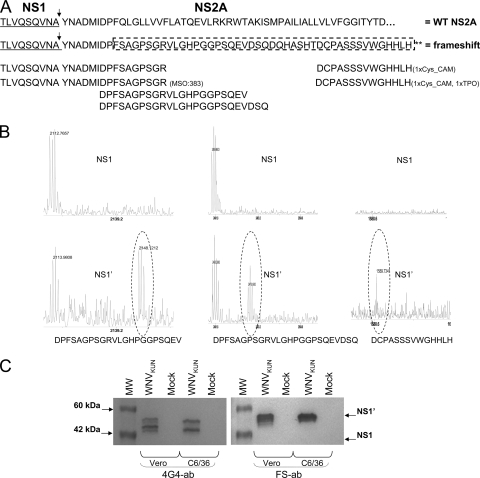
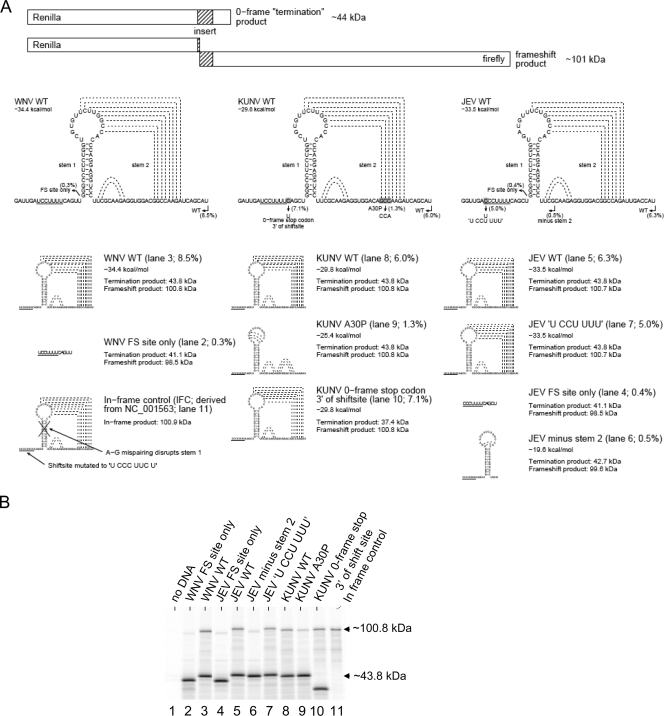
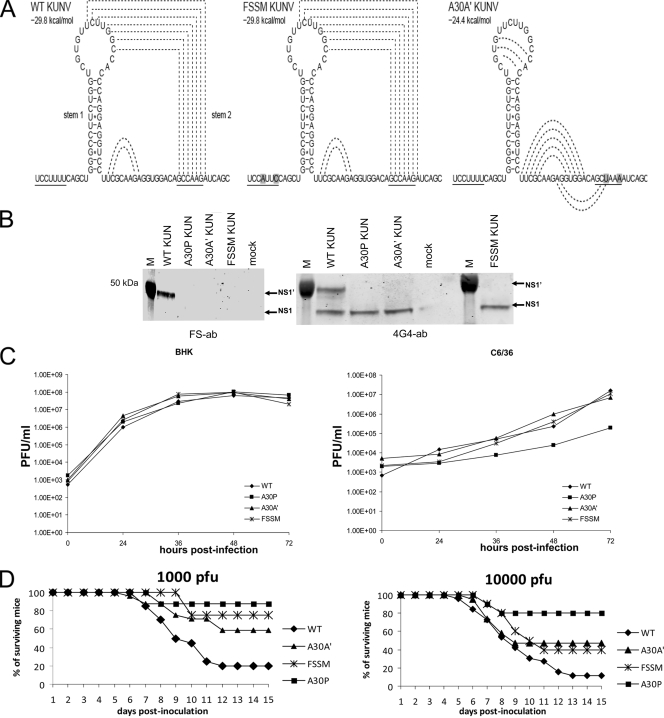
Flavivirus NS1 is a nonstructural protein involved in virus replication and regulation of the innate immune response. Interestingly, a larger NS1-related protein, NS1', is often detected during infection with the members of the Japanese encephalitis virus serogroup of flaviviruses. However, how NS1' is made and what role it performs in the viral life cycle have not been determined. Here we provide experimental evidence that NS1' is the product of a -1 ribosomal frameshift event that occurs at a conserved slippery heptanucleotide motif located near the beginning of the NS2A gene and is stimulated by a downstream RNA pseudoknot structure. Using site-directed mutagenesis of these sequence elements in an infectious clone of the Kunjin subtype of West Nile virus, we demonstrate that NS1' plays a role in viral neuroinvasiveness.
Figures
References
-
- Blitvich, B. J., J. S. Mackenzie, R. J. Coelen, M. J. Howard, and R. A. Hall. 1995. A novel complex formed between the flavivirus E and NS1 proteins: analysis of its structure and function. Arch. Virol. 140:145-156. - PubMed
-
- Blitvich, B. J., D. Scanlon, B. J. Shiell, J. S. Mackenzie, and R. A. Hall. 1999. Identification and analysis of truncated and elongated species of the flavivirus NS1 protein. Virus Res. 60:67-79. - PubMed
-
- Chen, L. K., C. L. Liao, C. G. Lin, S. C. Lai, C. I. Liu, S. H. Ma, Y. Y. Huang, and Y. L. Lin. 1996. Persistence of Japanese encephalitis virus is associated with abnormal expression of the nonstructural protein NS1 in host cells. Virology 217:220-229. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
