

Polar residues in the second transmembrane domain of the rat P2X2 receptor that affect spontaneous gating, unitary conductance, and rectification
- PMID: 19906973
- PMCID: PMC2804292
- DOI: 10.1523/JNEUROSCI.4403-09.2009
Polar residues in the second transmembrane domain of the rat P2X2 receptor that affect spontaneous gating, unitary conductance, and rectification
Abstract
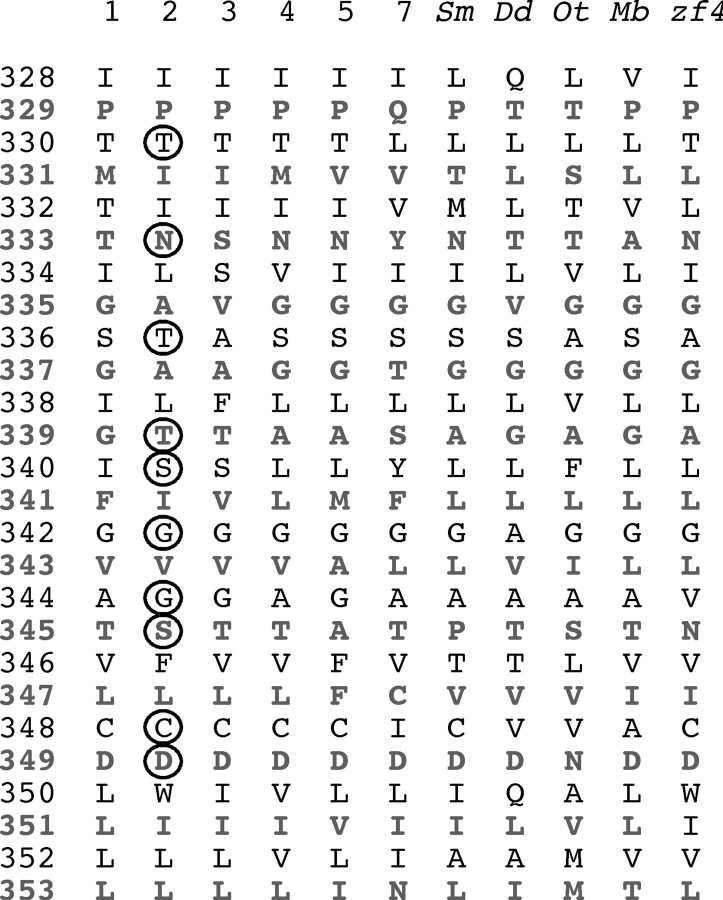
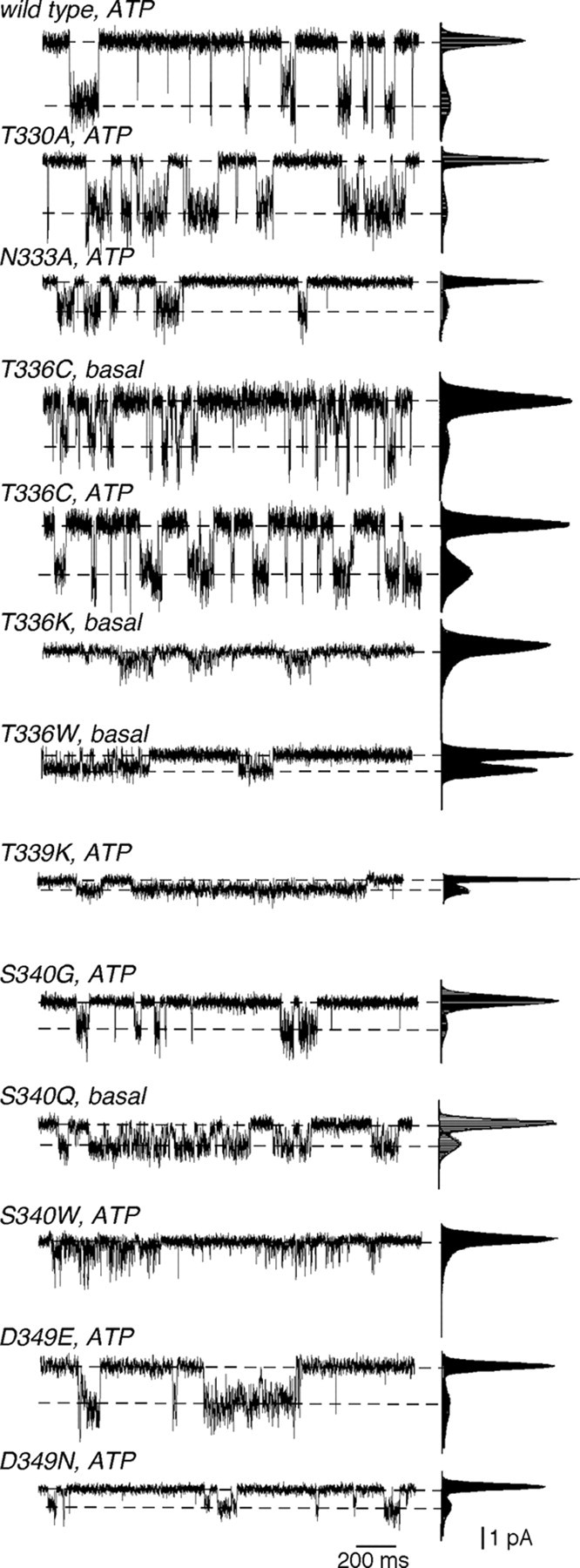
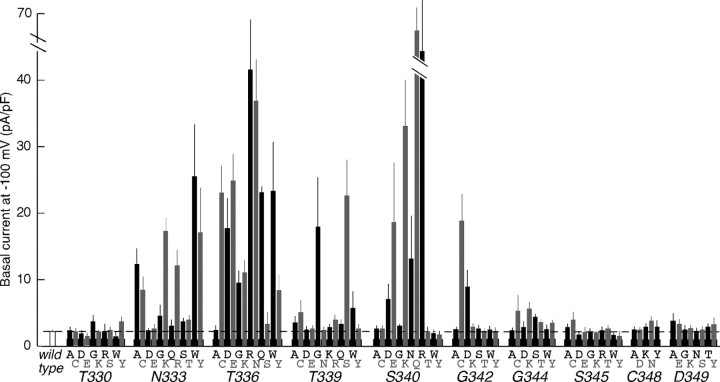
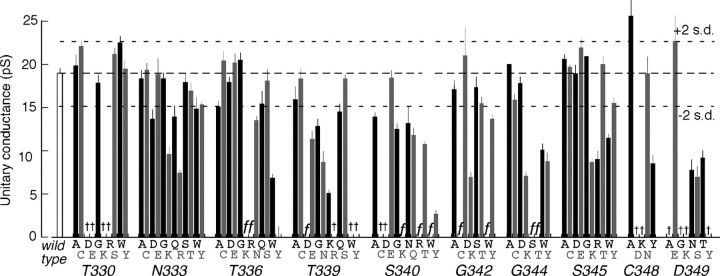
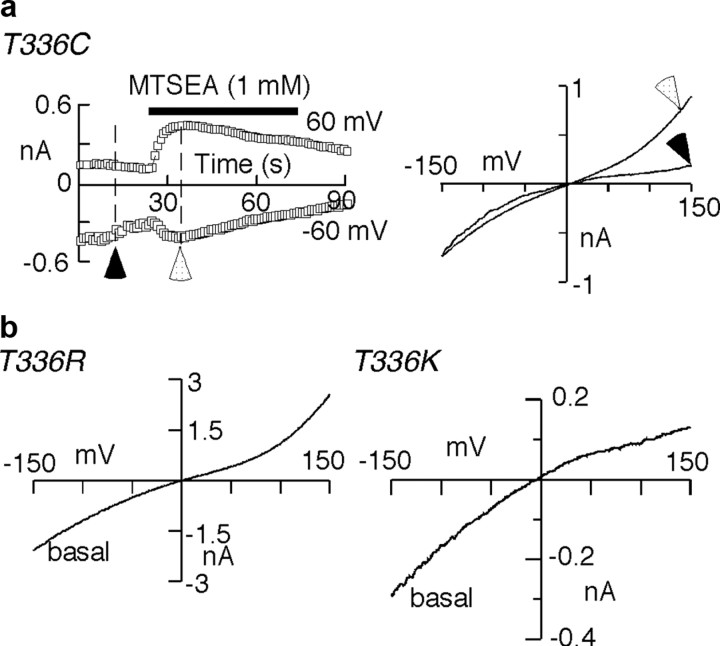
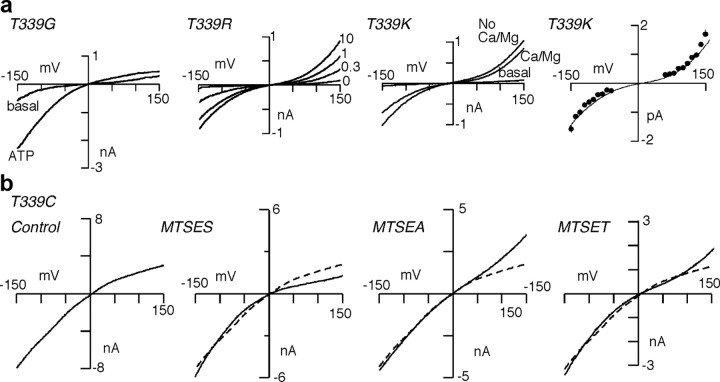
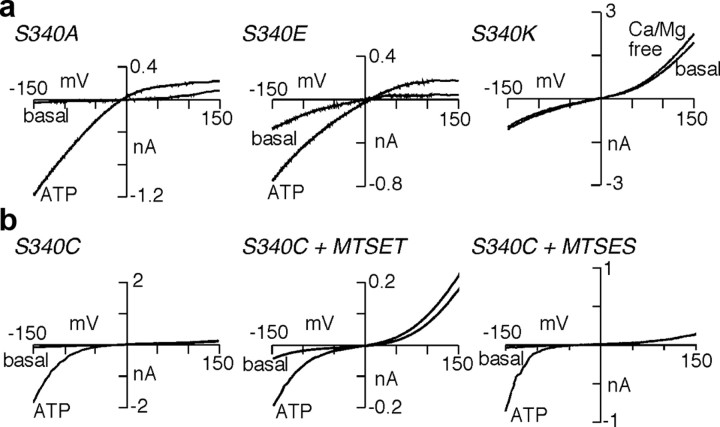
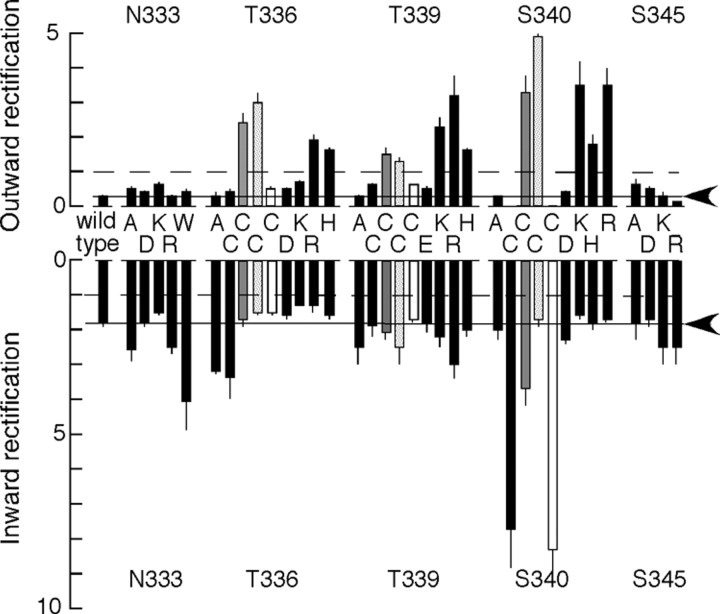
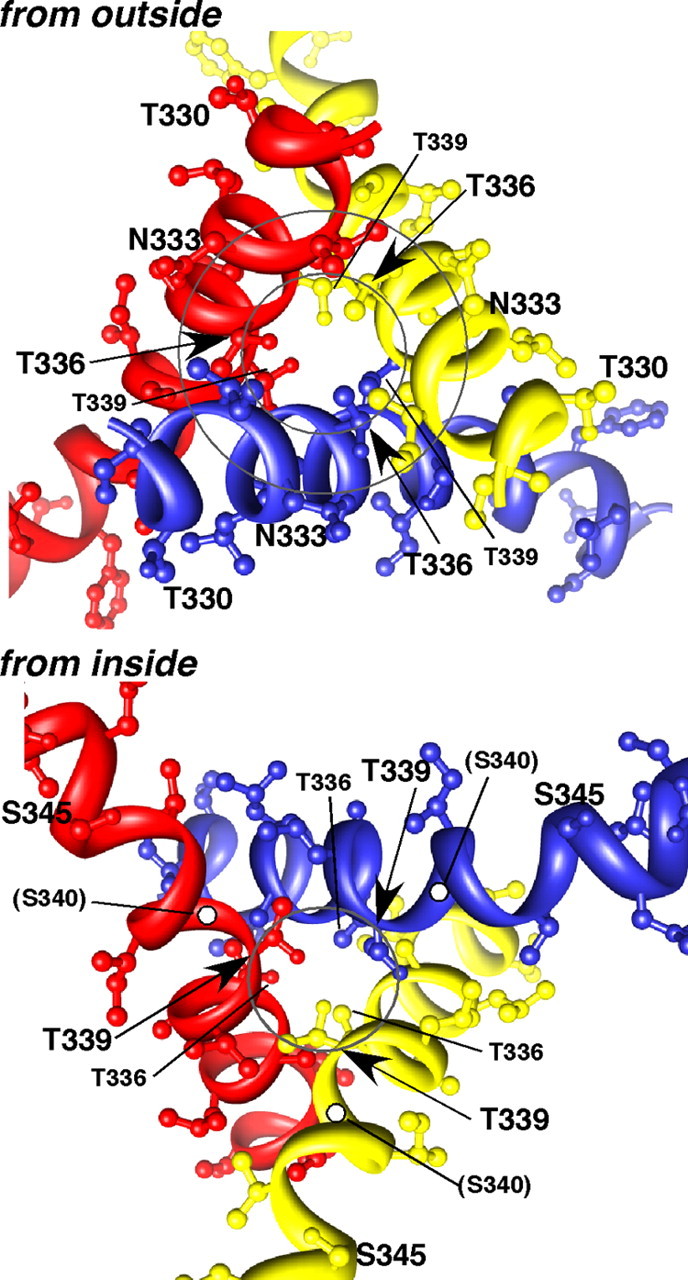
Membrane ion channels activated by extracellular ATP (P2X receptors) are widely distributed in the nervous system. Their molecular architecture is fundamentally distinct from that of the nicotinic or glutamate receptor families. We have measured single-channel currents, spontaneous gating, and rectification of rat P2X2 receptor in which polar and charged residues of the second transmembrane domain (TM2) were systematically probed by mutagenesis. The results suggest that Asn(333) and Asp(349) lie respectively in external and internal vestibules. Substitutions at Asn(333), Thr(336), and Ser(340) were particularly likely to cause spontaneously active channels. At Thr(336), Thr(339), and Ser(340), the introduction of positive charge (Arg, Lys, or His, or Cys followed by treatment with 2-aminoethyl methanethiosulphonate) greatly enhanced outward currents, suggesting that side-chains of these three residues are exposed in the permeation pathway of the open channel. These functional findings are interpreted in the context of the recently reported 3.1 A crystal structure of the zebrafish P2X4.1 receptor in the closed state. They imply that the gate is formed by residues Asn(333) to Thr(339) and that channel opening involves a counter-clockwise rotation and separation of the TM2 helices.
Figures
References
-
- Brake AJ, Wagenbach MJ, Julius D. New structural motif for ligand-gated ion channels defined by an ionotropic ATP receptor. Nature. 1994;371:519–523. - PubMed
-
- Clyne JD, Brown TC, Hume RI. Expression level dependent changes in the properties of P2X2 receptors. Neuropharmacology. 2003;44:403–412. - PubMed
-
- Ding S, Sachs F. Ion permeation and block of P2X2 purinoceptors: single channel recordings. J Membr Biol. 1999b;172:215–223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases