

Nonoptimal microbial response to antibiotics underlies suppressive drug interactions
- PMID: 19914165
- PMCID: PMC2838386
- DOI: 10.1016/j.cell.2009.10.025
Nonoptimal microbial response to antibiotics underlies suppressive drug interactions
Abstract
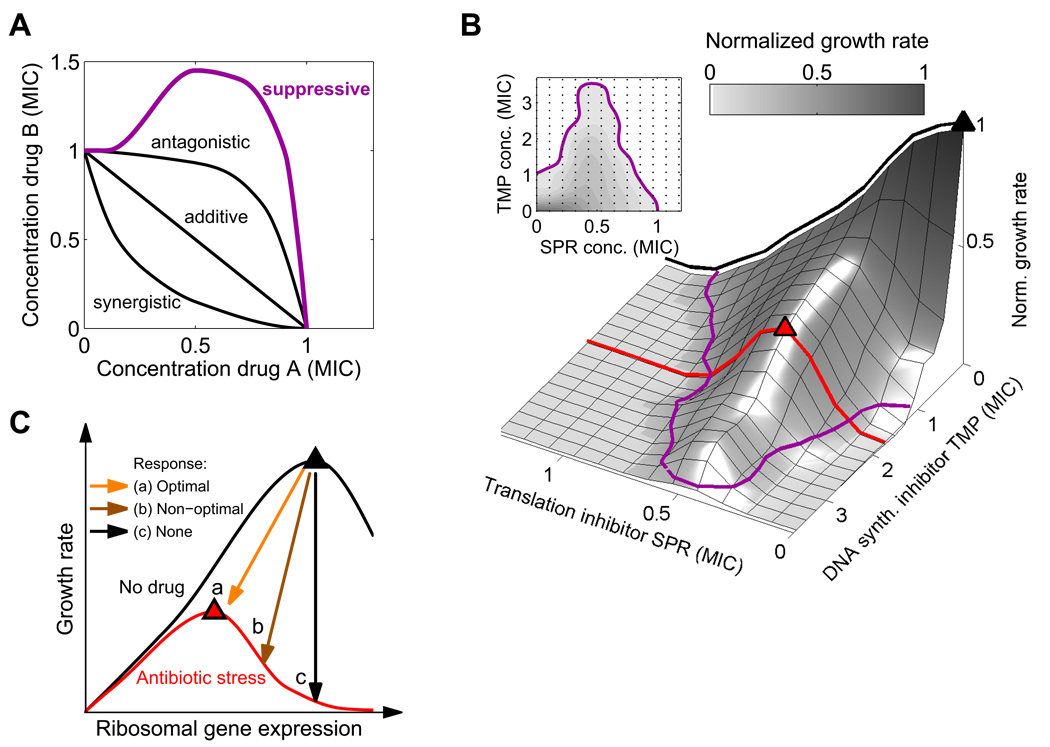
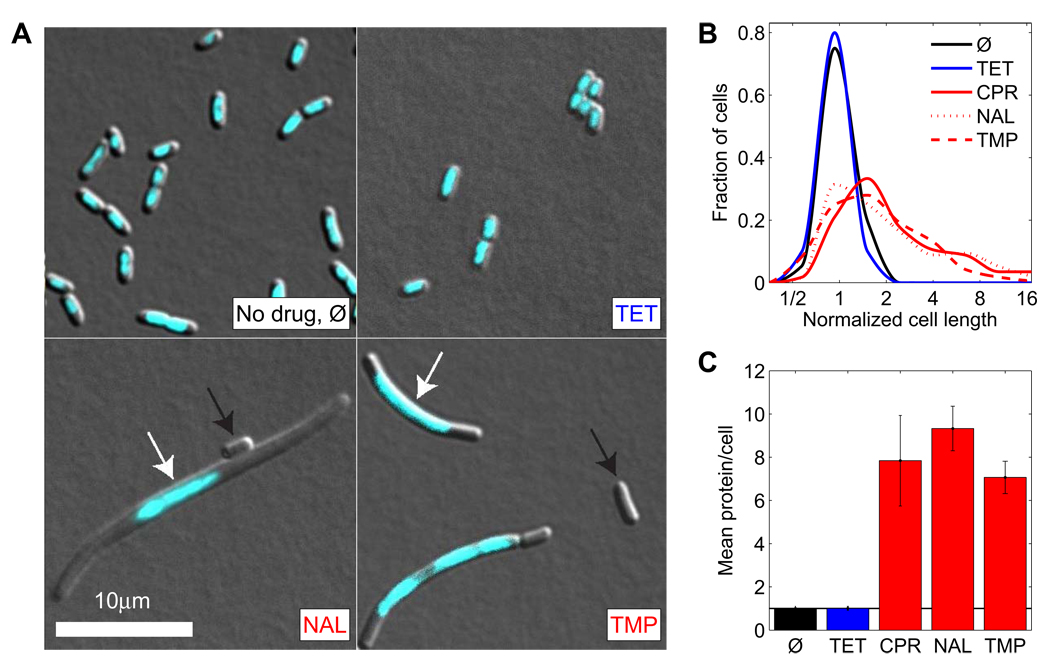
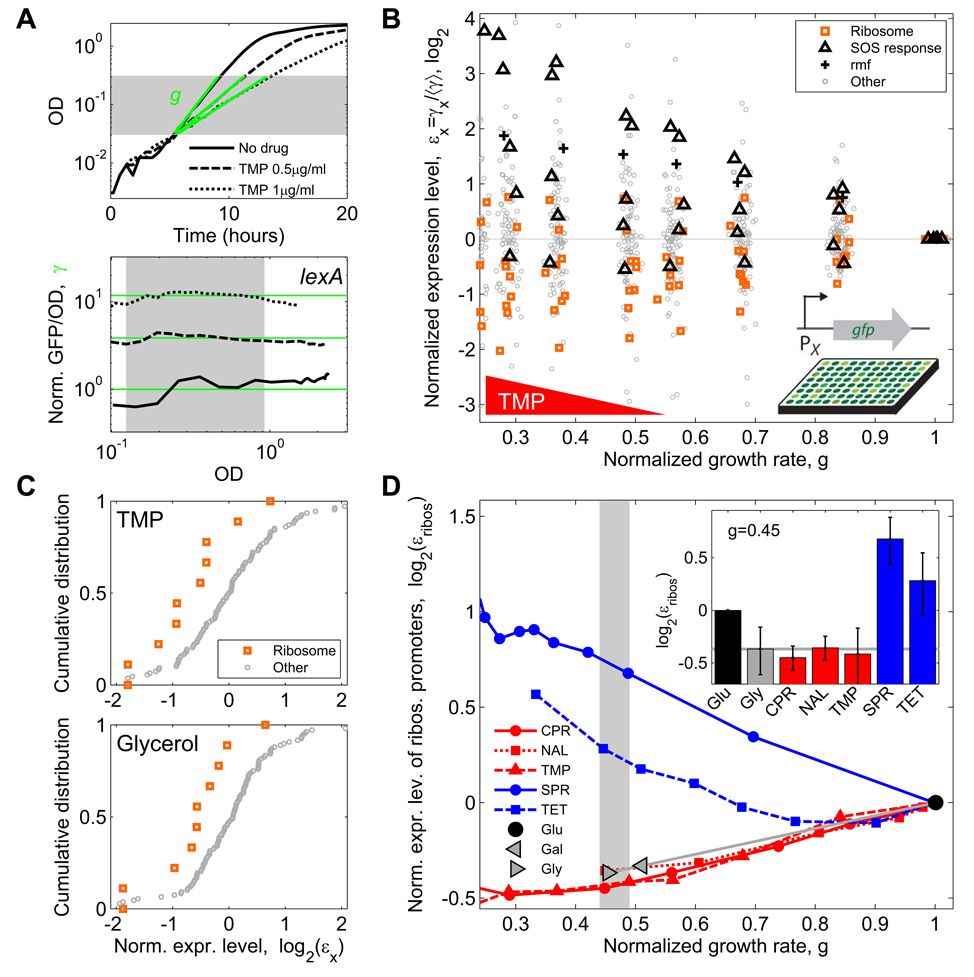
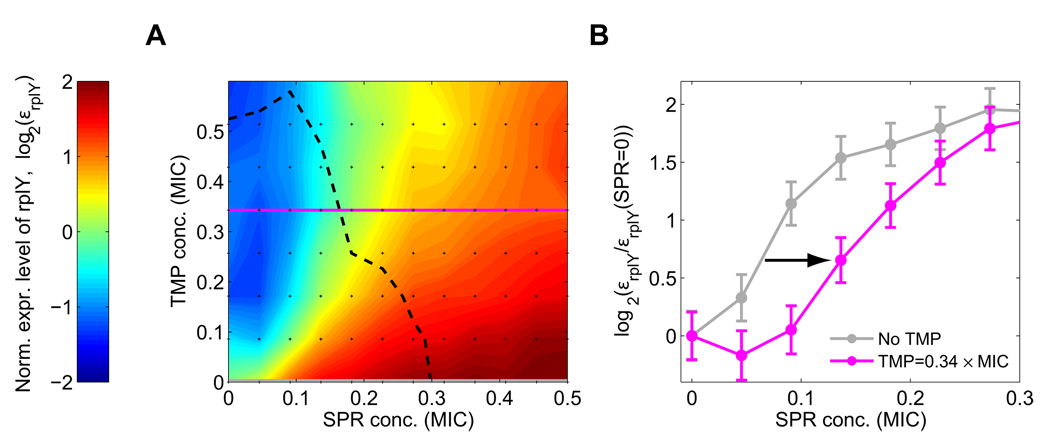
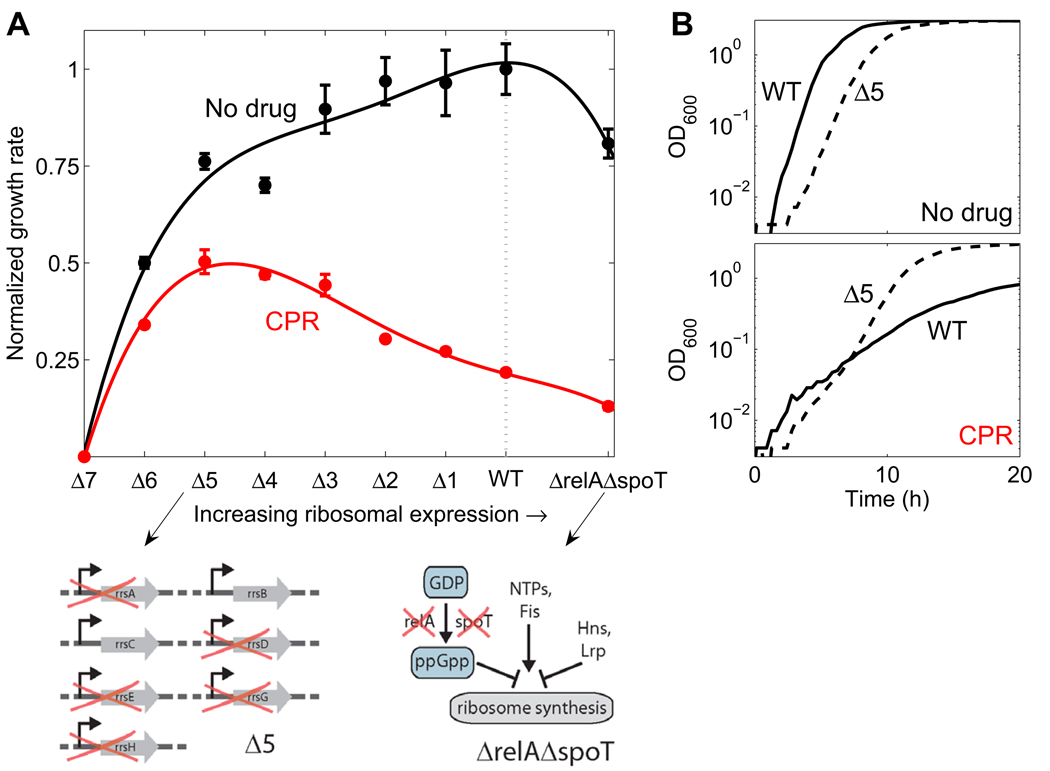
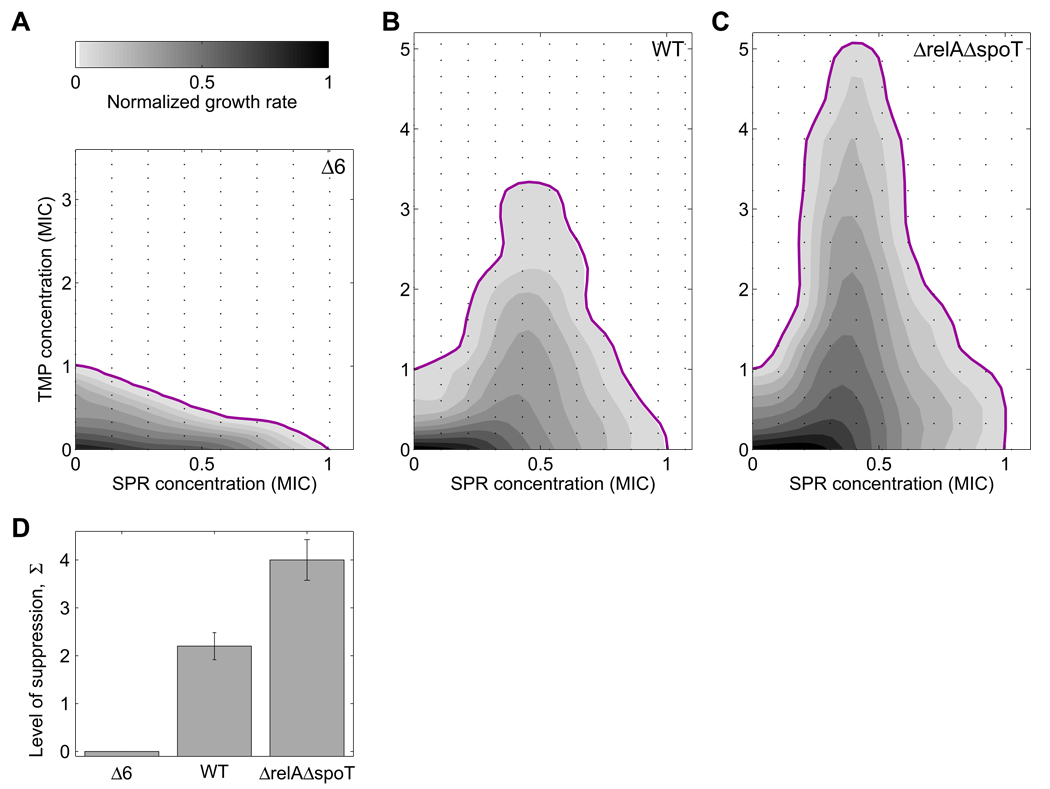
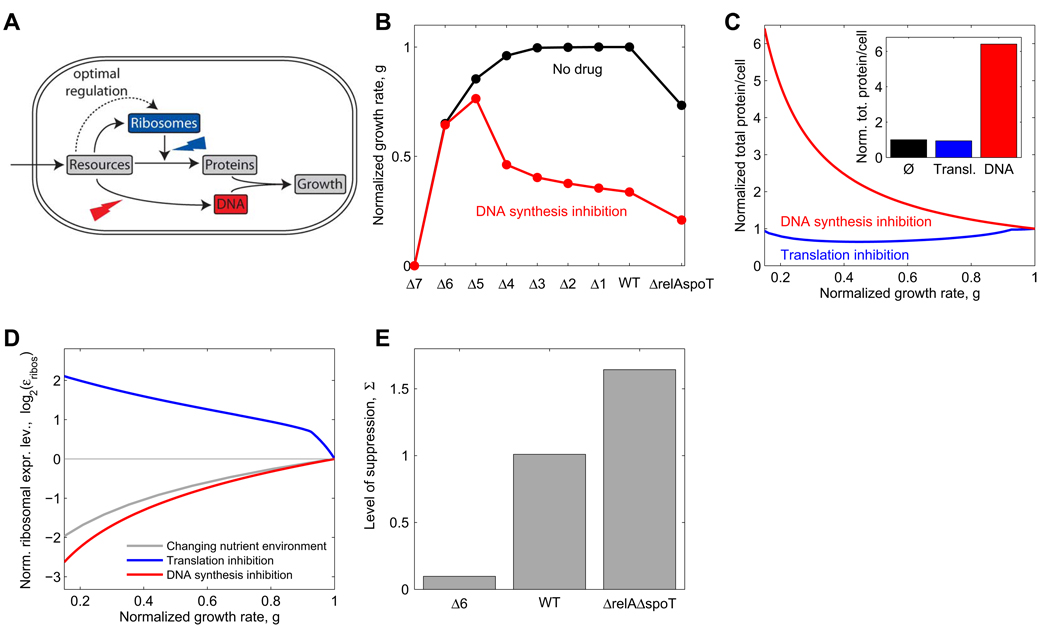
Suppressive drug interactions, in which one antibiotic can actually help bacterial cells to grow faster in the presence of another, occur between protein and DNA synthesis inhibitors. Here, we show that this suppression results from nonoptimal regulation of ribosomal genes in the presence of DNA stress. Using GFP-tagged transcription reporters in Escherichia coli, we find that ribosomal genes are not directly regulated by DNA stress, leading to an imbalance between cellular DNA and protein content. To test whether ribosomal gene expression under DNA stress is nonoptimal for growth rate, we sequentially deleted up to six of the seven ribosomal RNA operons. These synthetic manipulations of ribosomal gene expression correct the protein-DNA imbalance, lead to improved survival and growth, and completely remove the suppressive drug interaction. A simple mathematical model explains the nonoptimal regulation in different nutrient environments. These results reveal the genetic mechanism underlying an important class of suppressive drug interactions.
Figures
References
-
- Balaban NQ, Merrin J, Chait R, Kowalik L, Leibler S. Bacterial persistence as a phenotypic switch. Science. 2004;305:1622–1625. - PubMed
-
- Barker MM, Gaal T, Gourse RL. Mechanism of regulation of transcription initiation by ppGpp. II. Models for positive control based on properties of RNAP mutants and competition for RNAP. J Mol Biol. 2001;305:689–702. - PubMed
-
- Bliss CI. The toxicity of poisons applied jointly. Annals of Applied Biology. 1939;26:585–615.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
