

Transgenic microRNA inhibition with spatiotemporal specificity in intact organisms
- PMID: 19915559
- PMCID: PMC3183579
- DOI: 10.1038/nmeth.1402
Transgenic microRNA inhibition with spatiotemporal specificity in intact organisms
Abstract
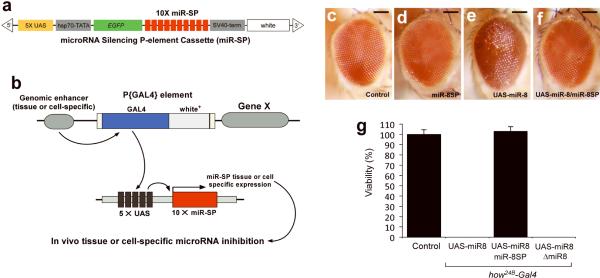
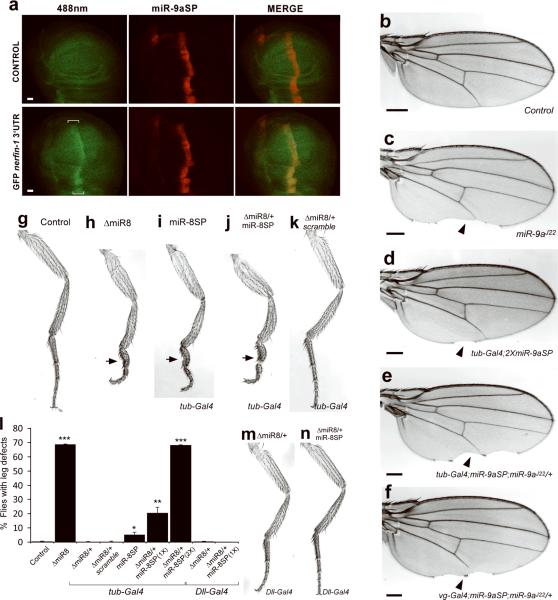
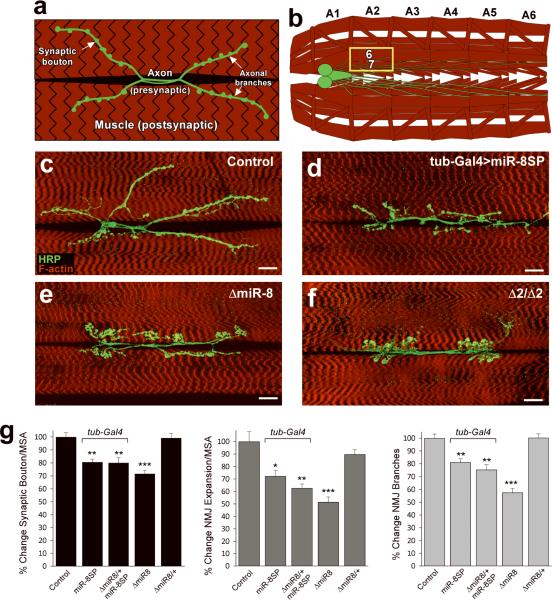
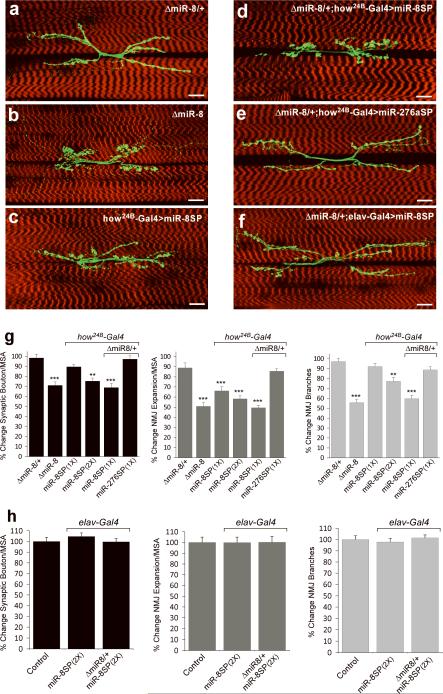
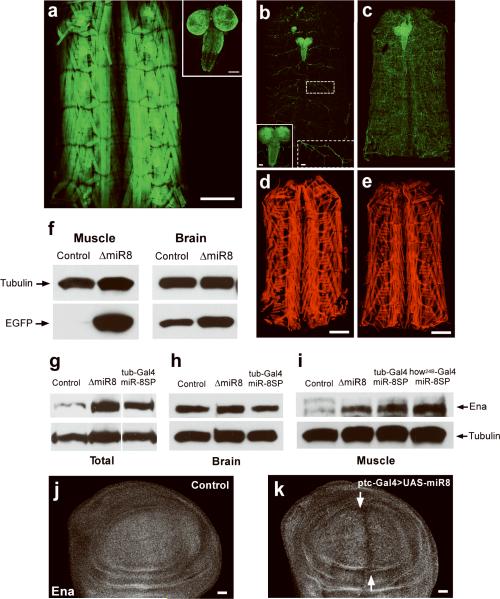
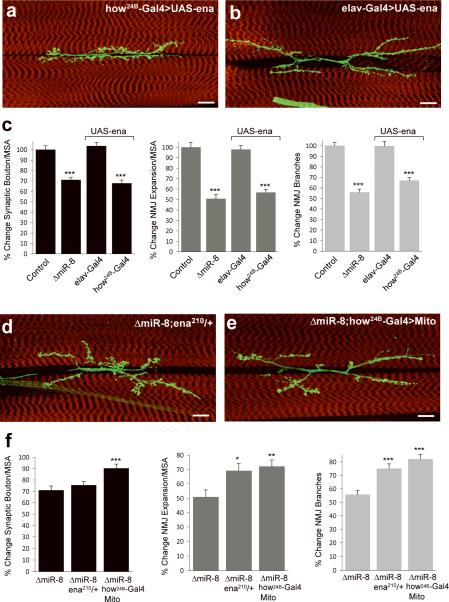
MicroRNAs are important regulators of gene expression, yet the functional outputs of most microRNA-target interactions remain elusive. Here we introduce the Drosophila melanogaster microRNA sponge (miR-SP) as a powerful transgenic technology to dissect the function of every microRNA with precise spatiotemporal resolution. miR-SPs can be used to characterize tissue-specific microRNA loss-of-function phenotypes, define the spatial regulation of their effectors and uncover interactions between microRNAs and other genes. Using themiR-SP system, we identified an essential role of the conserved microRNA miR-8, in neuromuscular junction formation. Tissue-specific silencing revealed that postsynaptic activity of miR-8 is important for normal neuromuscular junction morphogenesis. Given that miR-SPs rely on a bipartite modular expression system, they could be used to elucidate the endogenous function of microRNAs in any species in which conditional expression can be achieved.
Figures
Comment in
-
Use of microRNA sponges to explore tissue-specific microRNA functions in vivo.Nat Methods. 2009 Dec;6(12):873-4. doi: 10.1038/nmeth1209-873. Nat Methods. 2009. PMID: 19935840
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
