

Attention improves performance primarily by reducing interneuronal correlations
- PMID: 19915566
- PMCID: PMC2820564
- DOI: 10.1038/nn.2439
Attention improves performance primarily by reducing interneuronal correlations
Abstract
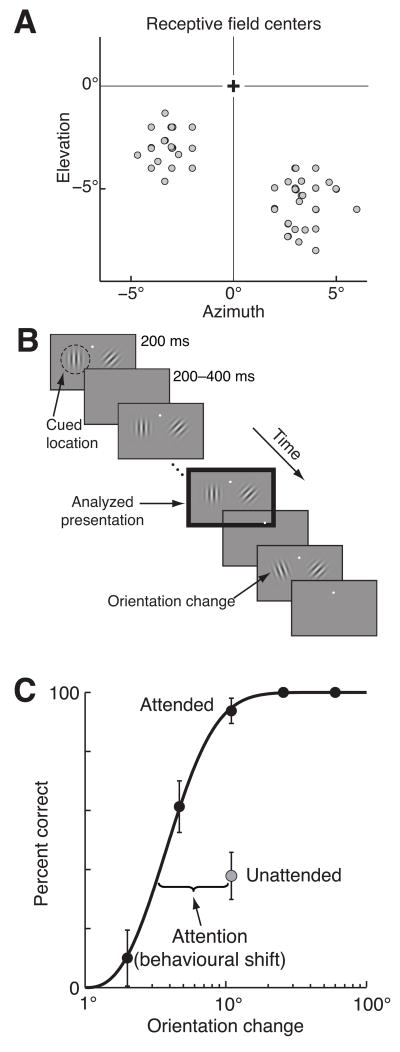
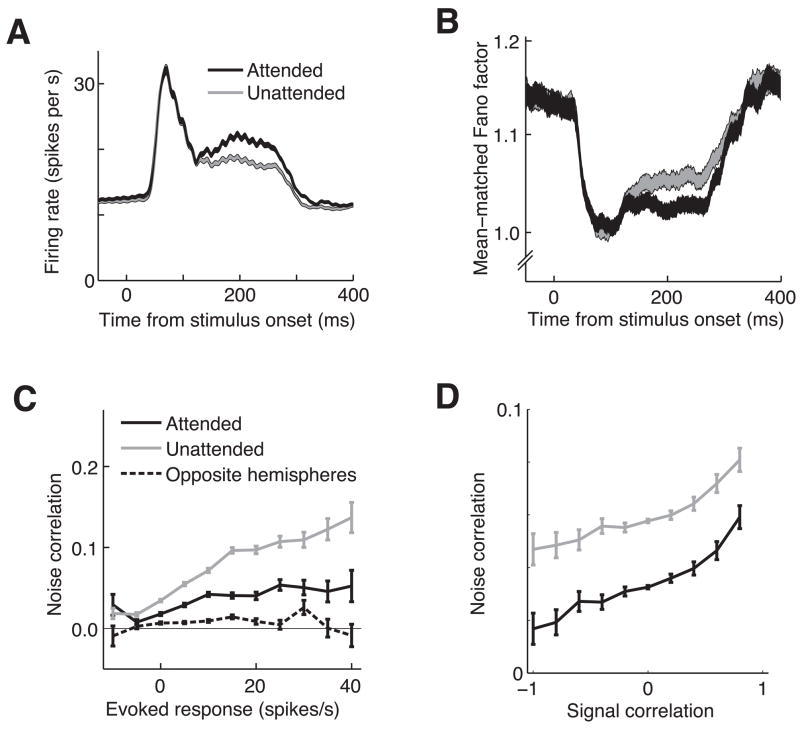
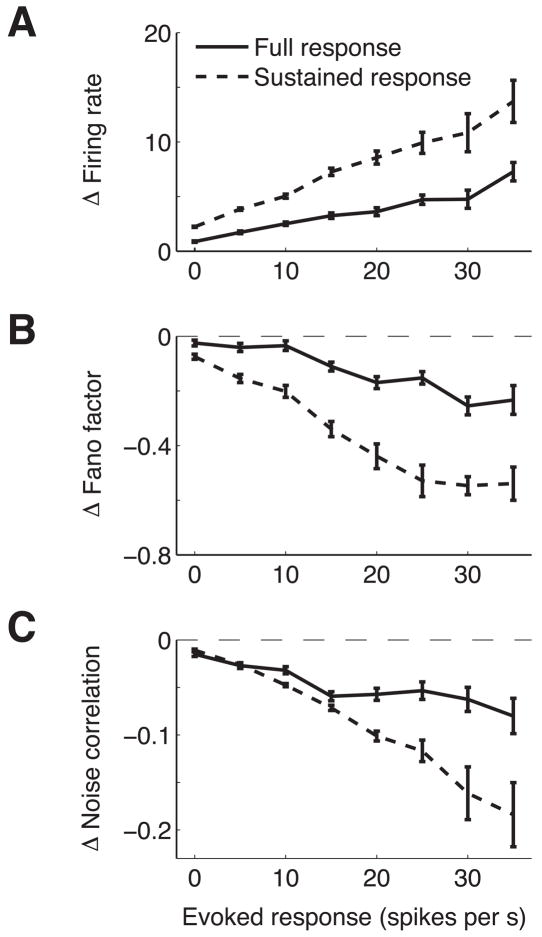
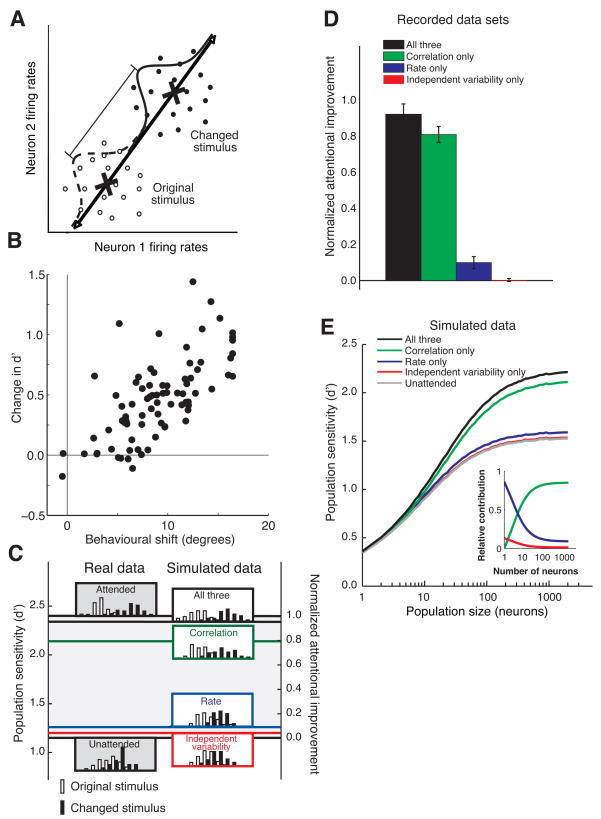
Visual attention can improve behavioral performance by allowing observers to focus on the important information in a complex scene. Attention also typically increases the firing rates of cortical sensory neurons. Rate increases improve the signal-to-noise ratio of individual neurons, and this improvement has been assumed to underlie attention-related improvements in behavior. We recorded dozens of neurons simultaneously in visual area V4 and found that changes in single neurons accounted for only a small fraction of the improvement in the sensitivity of the population. Instead, over 80% of the attentional improvement in the population signal was caused by decreases in the correlations between the trial-to-trial fluctuations in the responses of pairs of neurons. These results suggest that the representation of sensory information in populations of neurons and the way attention affects the sensitivity of the population may only be understood by considering the interactions between neurons.
Figures
Similar articles
-
Attention Increases Spike Count Correlations between Visual Cortical Areas.J Neurosci. 2016 Jul 13;36(28):7523-34. doi: 10.1523/JNEUROSCI.0610-16.2016. J Neurosci. 2016. PMID: 27413161 Free PMC article.
-
When attention wanders: how uncontrolled fluctuations in attention affect performance.J Neurosci. 2011 Nov 2;31(44):15802-6. doi: 10.1523/JNEUROSCI.3063-11.2011. J Neurosci. 2011. PMID: 22049423 Free PMC article.
-
Differential attention-dependent response modulation across cell classes in macaque visual area V4.Neuron. 2007 Jul 5;55(1):131-41. doi: 10.1016/j.neuron.2007.06.018. Neuron. 2007. PMID: 17610822
-
Attentional modulation of visual processing.Annu Rev Neurosci. 2004;27:611-47. doi: 10.1146/annurev.neuro.26.041002.131039. Annu Rev Neurosci. 2004. PMID: 15217345 Review.
-
Attention and active vision.Vision Res. 2009 Jun;49(10):1233-48. doi: 10.1016/j.visres.2008.06.017. Epub 2008 Aug 3. Vision Res. 2009. PMID: 18627774 Free PMC article. Review.
Cited by
-
Rodent auditory perception: Critical band limitations and plasticity.Neuroscience. 2015 Jun 18;296:55-65. doi: 10.1016/j.neuroscience.2015.03.053. Epub 2015 Mar 28. Neuroscience. 2015. PMID: 25827498 Free PMC article. Review.
-
Individuals with autism spectrum disorder have altered visual encoding capacity.PLoS Biol. 2021 May 12;19(5):e3001215. doi: 10.1371/journal.pbio.3001215. eCollection 2021 May. PLoS Biol. 2021. PMID: 33979326 Free PMC article.
-
Measuring Fisher information accurately in correlated neural populations.PLoS Comput Biol. 2015 Jun 1;11(6):e1004218. doi: 10.1371/journal.pcbi.1004218. eCollection 2015 Jun. PLoS Comput Biol. 2015. PMID: 26030735 Free PMC article.
-
Short-term synaptic depression and stochastic vesicle dynamics reduce and shape neuronal correlations.J Neurophysiol. 2013 Jan;109(2):475-84. doi: 10.1152/jn.00733.2012. Epub 2012 Oct 31. J Neurophysiol. 2013. PMID: 23114215 Free PMC article.
-
The autism spectrum disorder risk gene NEXMIF over-synchronizes hippocampal CA1 network and alters neuronal coding.Front Neurosci. 2023 Oct 27;17:1277501. doi: 10.3389/fnins.2023.1277501. eCollection 2023. Front Neurosci. 2023. PMID: 37965217 Free PMC article.
References
-
- Yantis S, Serences JT. Cortical mechanisms of space-based and object-based attentional control. Curr Opin Neurobiol. 2003;13:187–193. - PubMed
-
- Assad JA. Neural coding of behavioural relevance in parietal cortex. Curr Opin Neurobiol. 2003;13:194–197. - PubMed
-
- Reynolds JH, Chelazzi L. Attentional modulation of visual processing. Annu Rev Neurosci. 2004;27:611–647. - PubMed
-
- Maunsell JH, Treue S. Feature-based attention in visual cortex. Trends Neurosci. 2006;29:317–322. - PubMed
-
- Tolhurst DJ, Movshon JA, Dean AF. The statistical reliability of signals in single neurons in cat and monkey visual cortex. Vision Res. 1983;23:775–785. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
