

The chaperonin TRiC blocks a huntingtin sequence element that promotes the conformational switch to aggregation
- PMID: 19915590
- PMCID: PMC2788664
- DOI: 10.1038/nsmb.1700
The chaperonin TRiC blocks a huntingtin sequence element that promotes the conformational switch to aggregation
Abstract
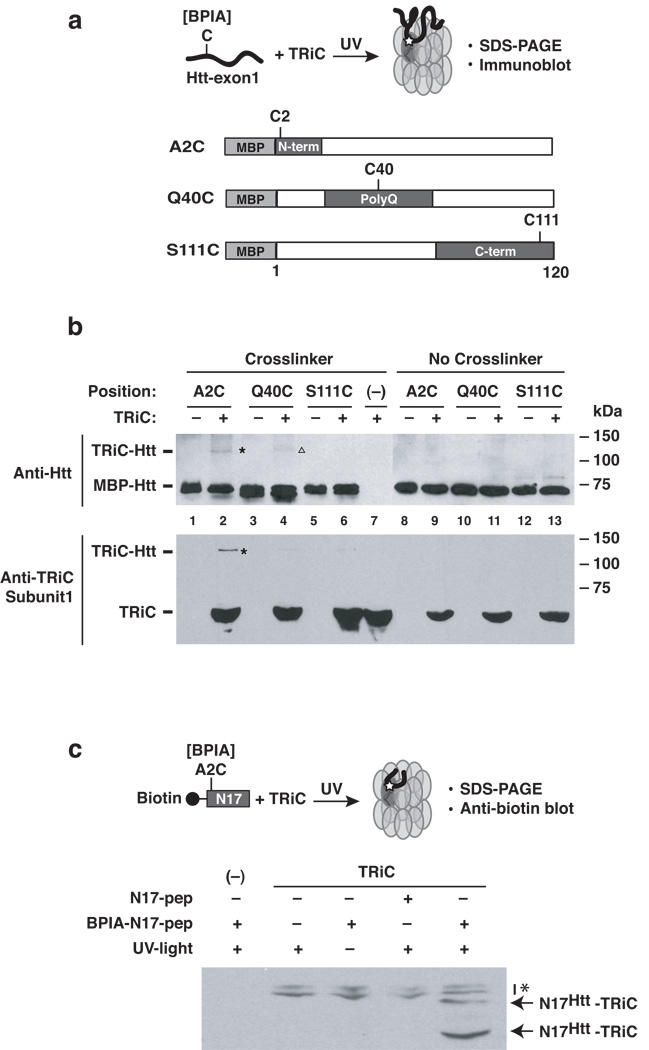
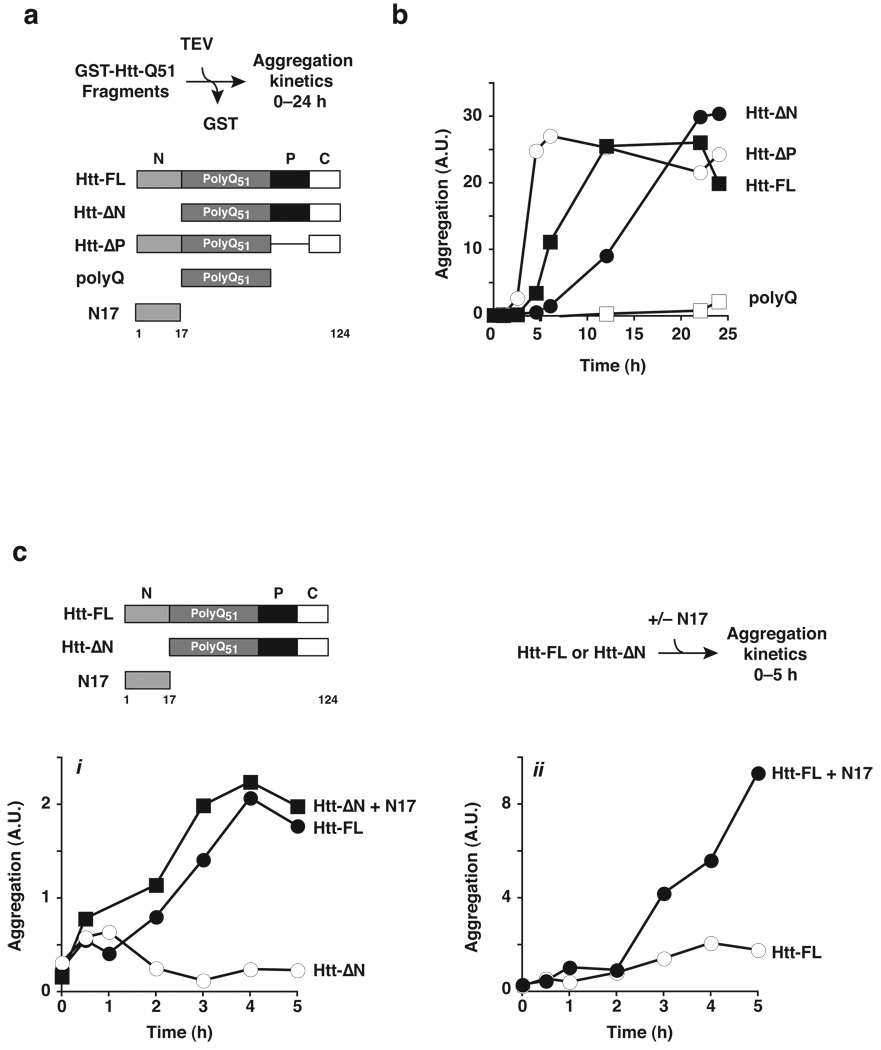
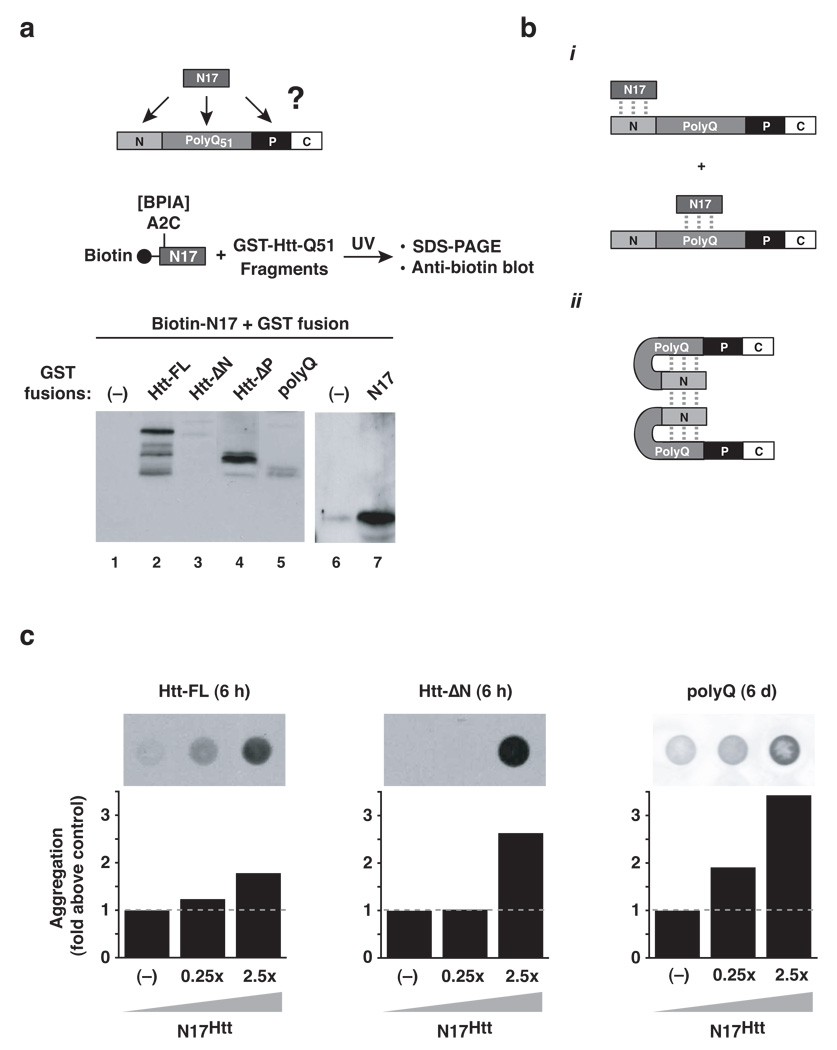
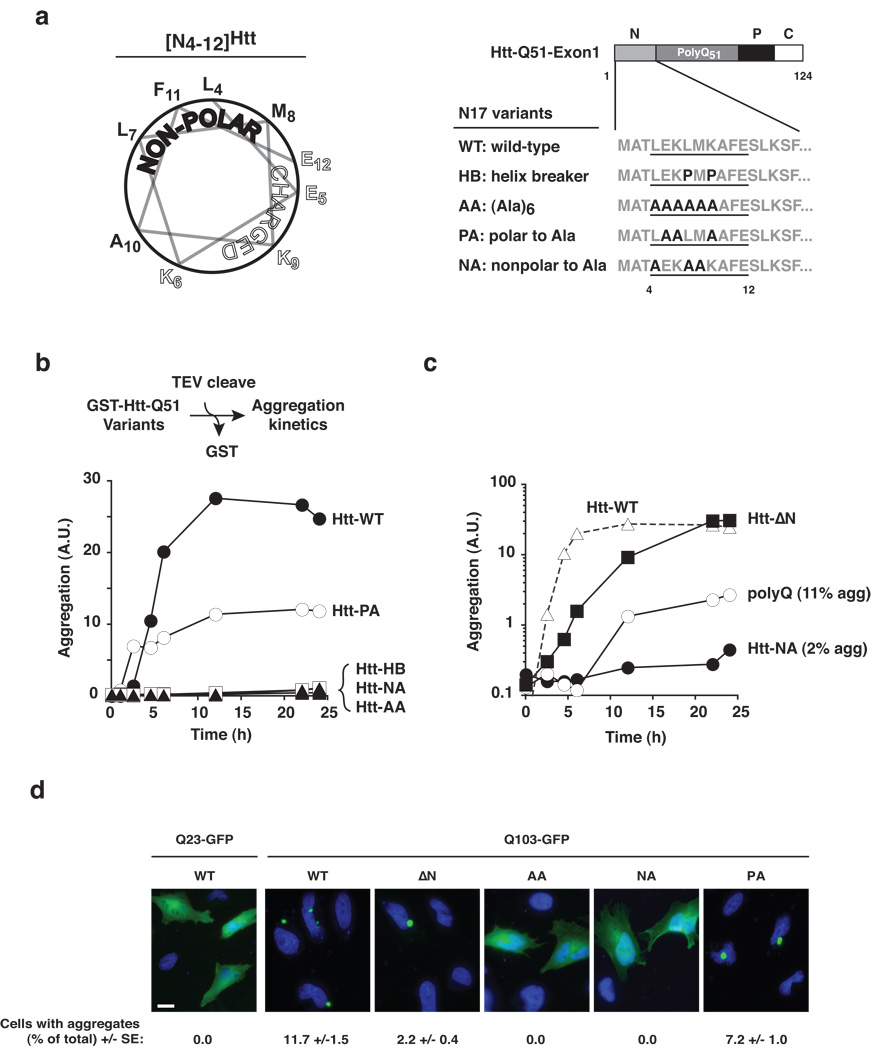
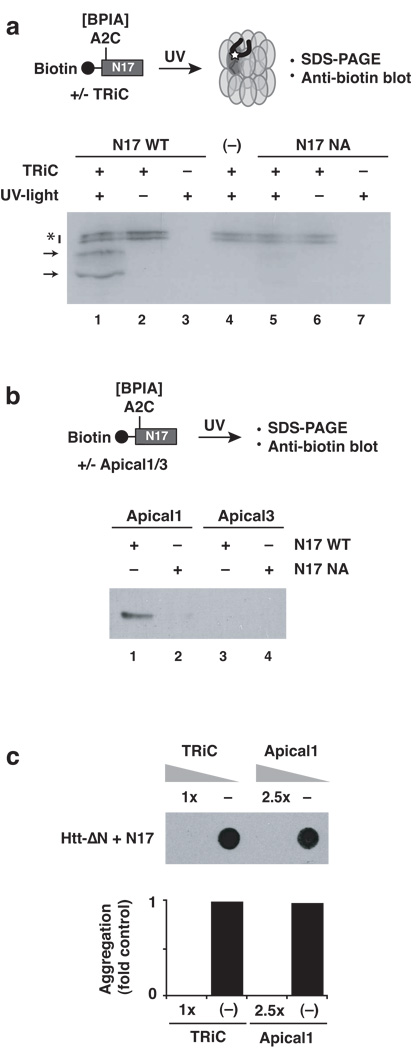
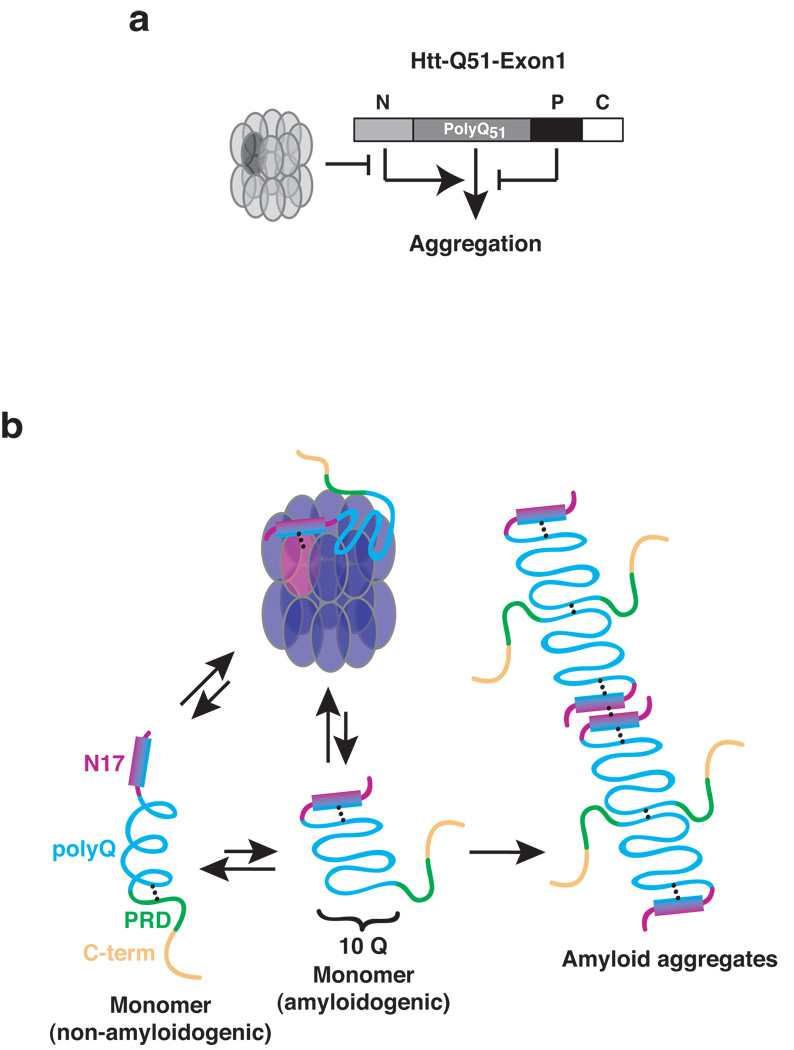
Aggregation of proteins containing polyglutamine (polyQ) expansions characterizes many neurodegenerative disorders, including Huntington's disease. Molecular chaperones modulate the aggregation and toxicity of the huntingtin (Htt) protein by an ill-defined mechanism. Here we determine how the chaperonin TRiC suppresses Htt aggregation. Unexpectedly, TRiC does not physically block the polyQ tract itself, but rather sequesters a short Htt sequence element, N-terminal to the polyQ tract, that promotes the amyloidogenic conformation. The residues of this element essential for rapid Htt aggregation are directly bound by TRiC. Our findings illustrate how molecular chaperones, which recognize hydrophobic determinants, can prevent aggregation of polar polyQ tracts associated with neurodegenerative diseases. The observation that short endogenous sequence elements can accelerate the switch of polyQ tracts to an amyloidogenic conformation provides a novel target for therapeutic strategies.
Figures
References
-
- Zoghbi HY, Orr HT. Glutamine repeats and neurodegeneration. Annu Rev Neurosci. 2000;23:217–247. - PubMed
-
- Soto C. Unfolding the role of protein misfolding in neurodegenerative diseases. Nat Rev Neurosci. 2003;4:49–60. - PubMed
-
- Chen S, Berthelier V, Yang W, Wetzel R. Polyglutamine aggregation behavior in vitro supports a recruitment mechanism of cytotoxicity. J Mol Biol. 2001;311:173–182. - PubMed
-
- Yang W, Dunlap JR, Andrews RB, Wetzel R. Aggregated polyglutamine peptides delivered to nuclei are toxic to mammalian cells. Hum Mol Genet. 2002;11:2905–2917. - PubMed
-
- Dobson CM. Principles of protein folding, misfolding and aggregation. Semin Cell Dev Biol. 2004;15:3–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
