

Ablation of arginylation in the mouse N-end rule pathway: loss of fat, higher metabolic rate, damaged spermatogenesis, and neurological perturbations
- PMID: 19915679
- PMCID: PMC2773024
- DOI: 10.1371/journal.pone.0007757
Ablation of arginylation in the mouse N-end rule pathway: loss of fat, higher metabolic rate, damaged spermatogenesis, and neurological perturbations
Abstract
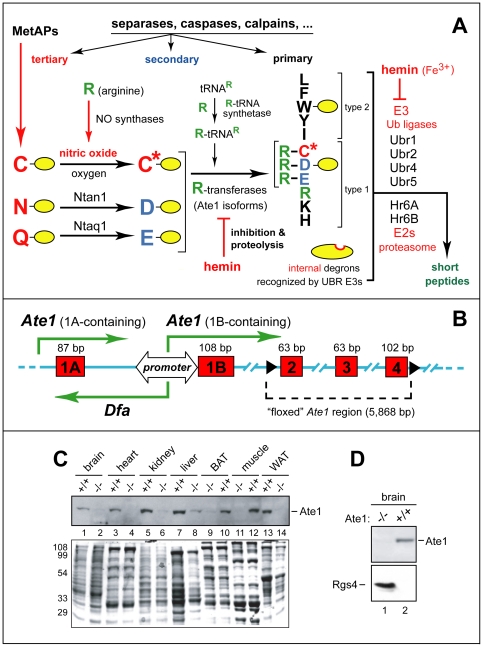
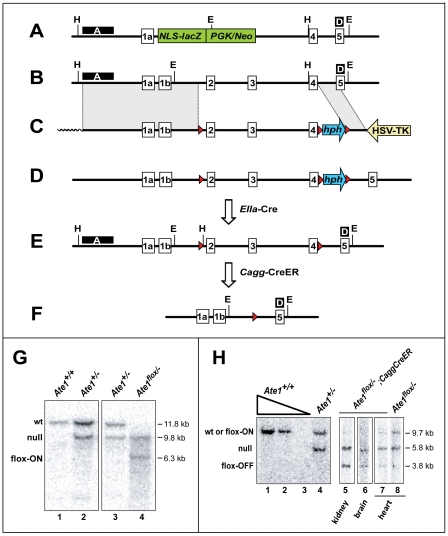
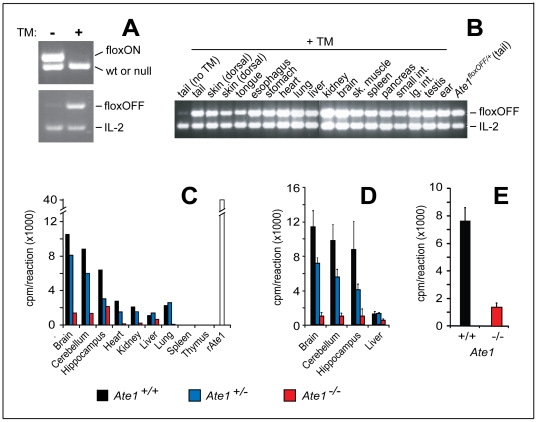
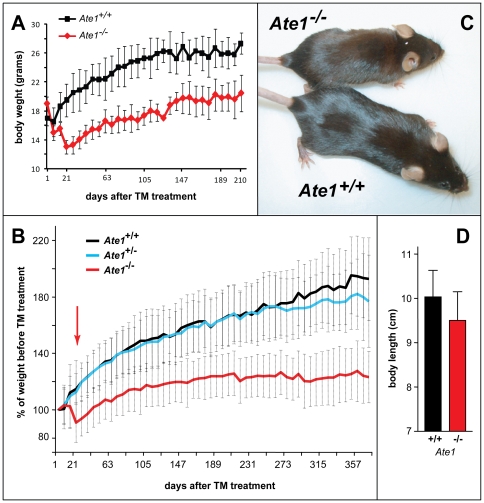
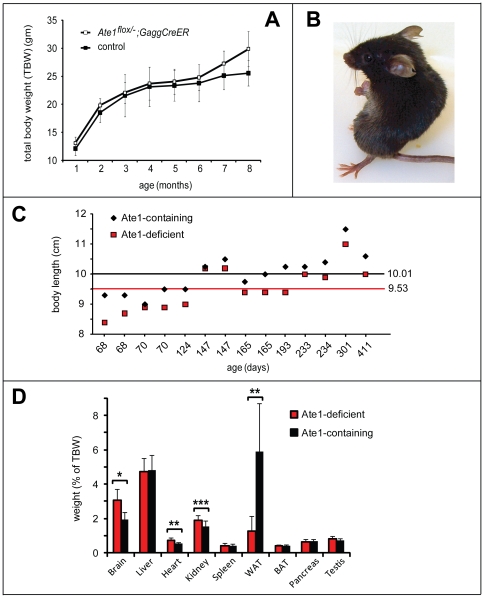
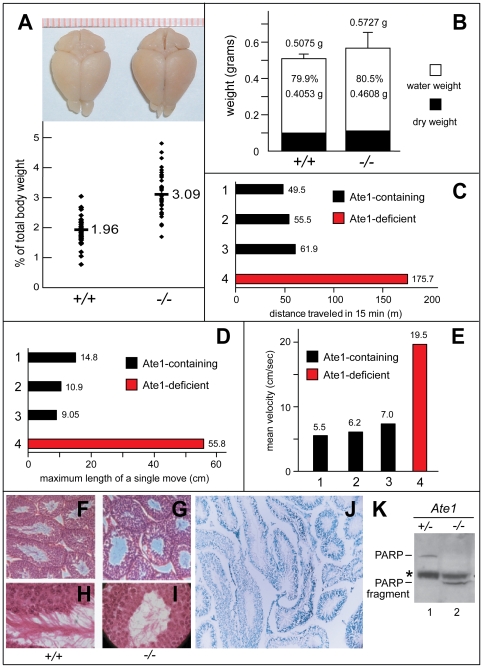
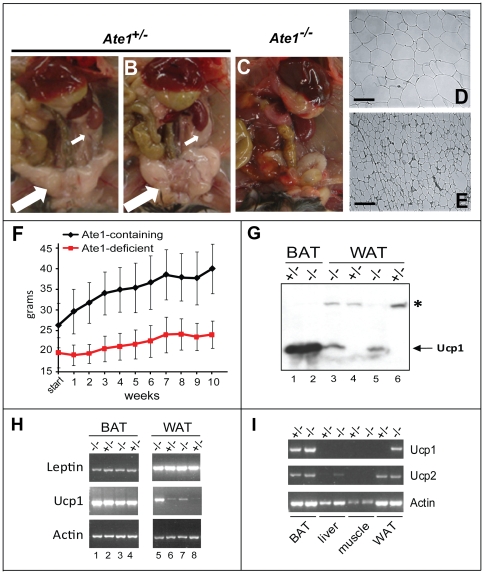
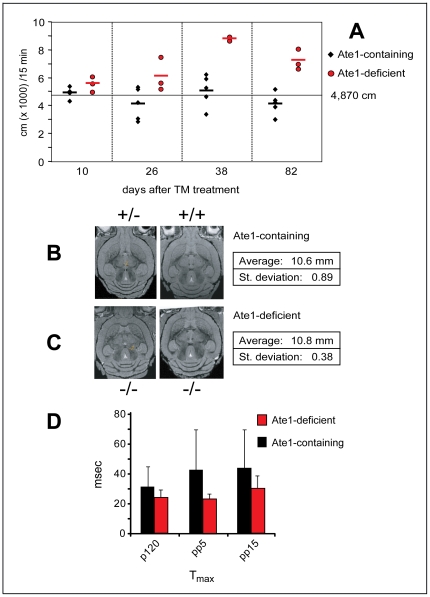
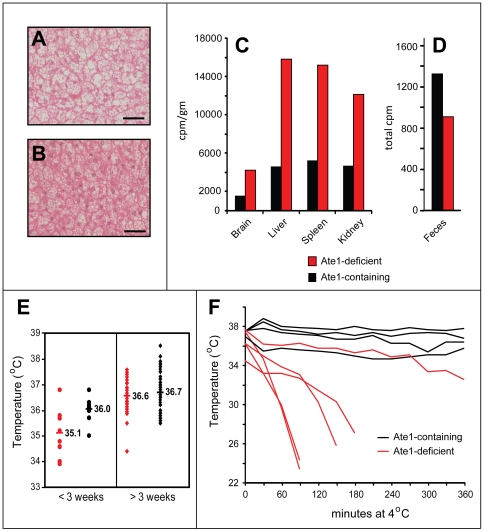
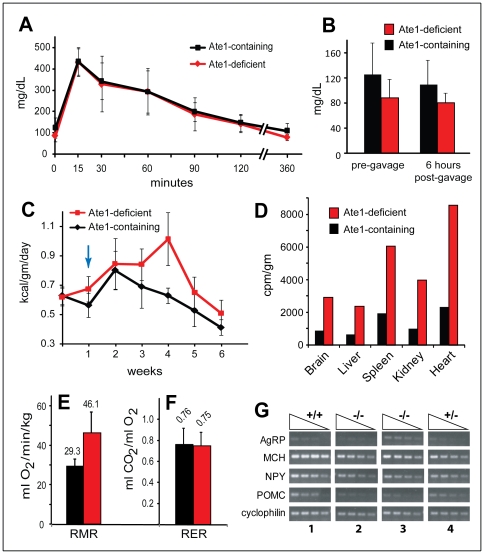
In the N-end rule pathway of protein degradation, the destabilizing activity of N-terminal Asp, Glu or (oxidized) Cys residues requires their conjugation to Arg, which is recognized directly by pathway's ubiquitin ligases. N-terminal arginylation is mediated by the Ate1 arginyltransferase, whose physiological substrates include the Rgs4, Rgs5 and Rgs16 regulators of G proteins. Here, we employed the Cre-lox technique to uncover new physiological functions of N-terminal arginylation in adult mice. We show that postnatal deletion of mouse Ate1 (its unconditional deletion is embryonic lethal) causes a rapid decrease of body weight and results in early death of approximately 15% of Ate1-deficient mice. Despite being hyperphagic, the surviving Ate1-deficient mice contain little visceral fat. They also exhibit an increased metabolic rate, ectopic induction of the Ucp1 uncoupling protein in white fat, and are resistant to diet-induced obesity. In addition, Ate1-deficient mice have enlarged brains, an enhanced startle response, are strikingly hyperkinetic, and are prone to seizures and kyphosis. Ate1-deficient males are also infertile, owing to defects in Ate1(-/-) spermatocytes. The remarkably broad range of specific biological processes that are shown here to be perturbed by the loss of N-terminal arginylation will make possible the dissection of regulatory circuits that involve Ate1 and either its known substrates, such as Rgs4, Rgs5 and Rgs16, or those currently unknown.
Conflict of interest statement
Figures
References
-
- Mogk A, Schmidt R, Bukau B. The N-end rule pathway of regulated proteolysis: prokaryotic and eukaryotic strategies. Trends Cell Biol. 2007;17:165–172. - PubMed
-
- Tasaki T, Kwon YT. The mammalian N-end rule pathway: new insights into its components and physiological roles. Trends Biochem Sci. 2007;32:520–528. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
