

Organization of ryanodine receptors, transverse tubules, and sodium-calcium exchanger in rat myocytes
- PMID: 19917219
- PMCID: PMC2776253
- DOI: 10.1016/j.bpj.2009.08.036
Organization of ryanodine receptors, transverse tubules, and sodium-calcium exchanger in rat myocytes
Abstract
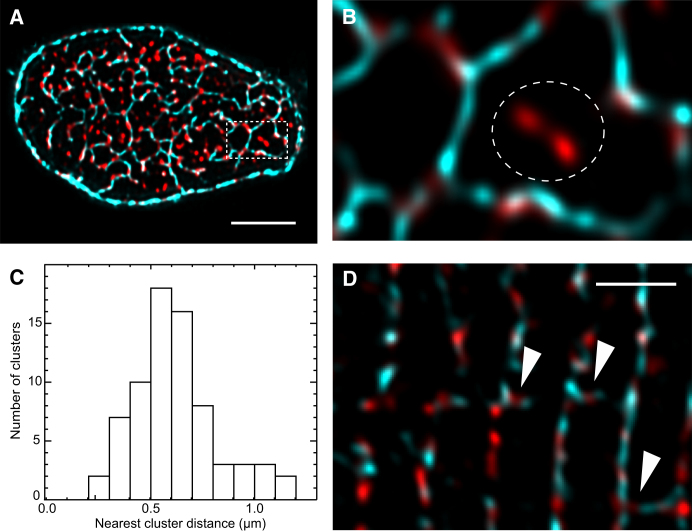
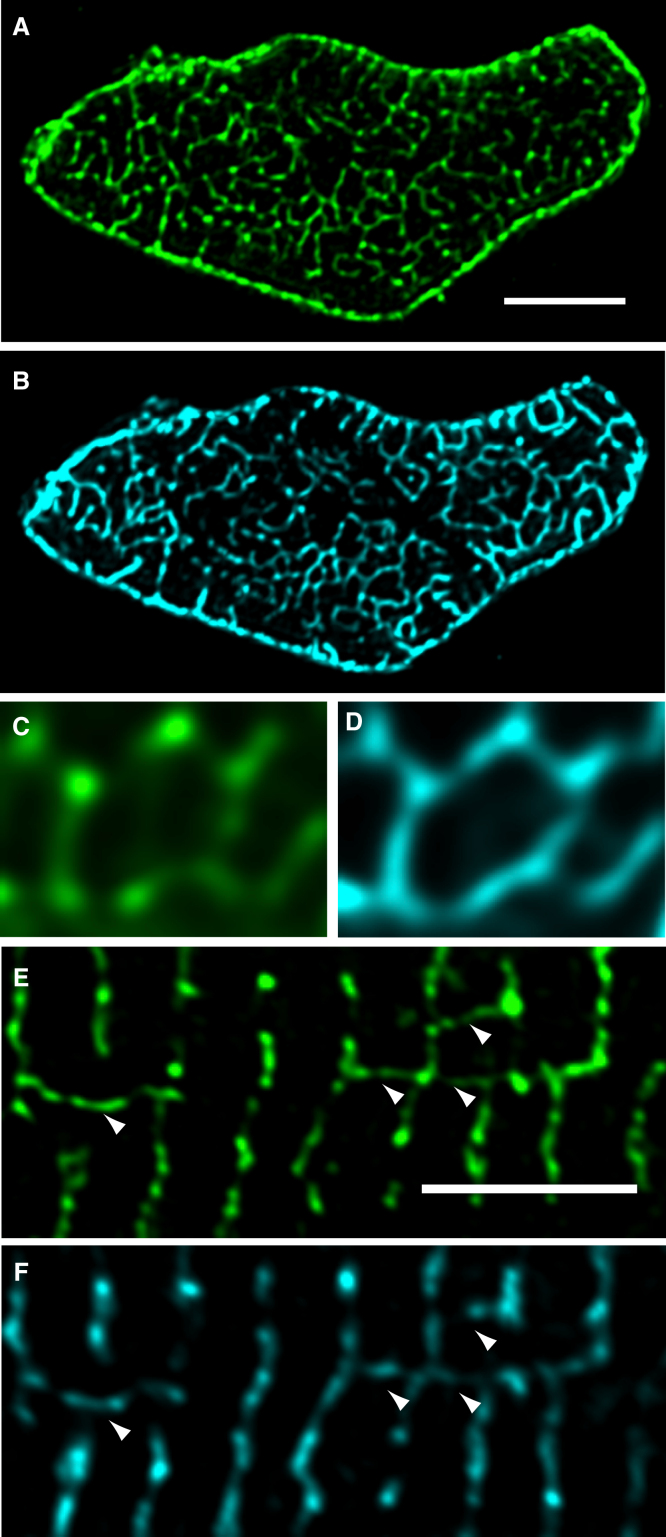
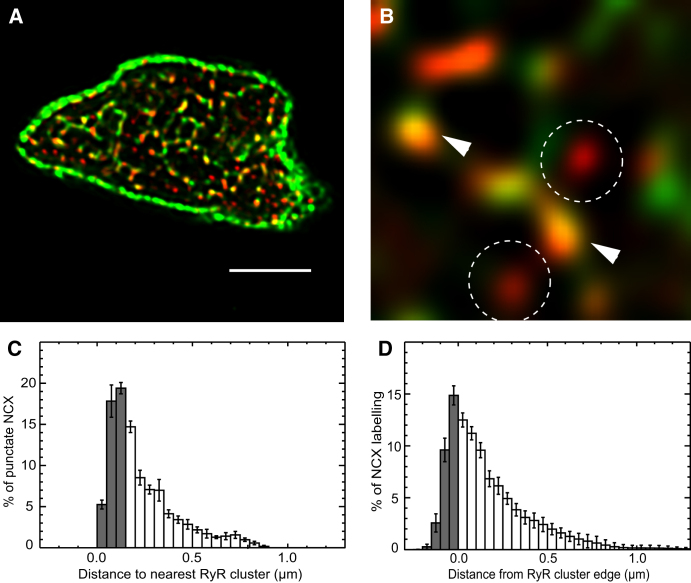
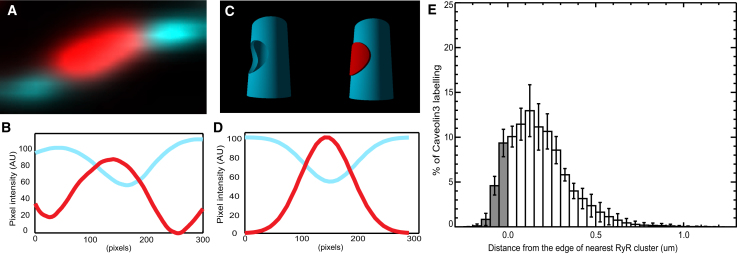
Confocal and total internal reflection fluorescence imaging was used to examine the distribution of caveolin-3, sodium-calcium exchange (NCX) and ryanodine receptors (RyRs) in rat ventricular myocytes. Transverse and longitudinal optical sectioning shows that NCX is distributed widely along the transverse and longitudinal tubular system (t-system). The NCX labeling consisted of both punctate and distributed components, which partially colocalize with RyRs (27%). Surface membrane labeling showed a similar pattern but the fraction of RyR clusters containing NCX label was decreased and no nonpunctate labeling was observed. Sixteen percent of RyRs were not colocalized with the t-system and 1.6% of RyRs were found on longitudinal elements of the t-system. The surface distribution of RyR labeling was not generally consistent with circular patches of RyRs. This suggests that previous estimates for the number of RyRs in a junction (based on circular close-packed arrays) need to be revised. The observed distribution of caveolin-3 labeling was consistent with its exclusion from RyR clusters. Distance maps for all colocalization pairs were calculated to give the distance between centroids of punctate labeling and edges for distributed components. The possible roles for punctate NCX labeling are discussed.
Figures


References
-
- Cannell M.B., Cheng H., Lederer W.J. The control of calcium release in the heart muscle. Science. 1995;268:1045–1049. - PubMed
-
- Soeller C., Cannell M.B. Examination of the transverse-tubular system in living cardiac rat myocytes by 2-photon microscopy and digital image-processing techniques. Circ. Res. 1999;84:266–275. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
