

Anthropomorphizing the mouse cardiac action potential via a novel dynamic clamp method
- PMID: 19917221
- PMCID: PMC2776258
- DOI: 10.1016/j.bpj.2009.09.002
Anthropomorphizing the mouse cardiac action potential via a novel dynamic clamp method
Abstract
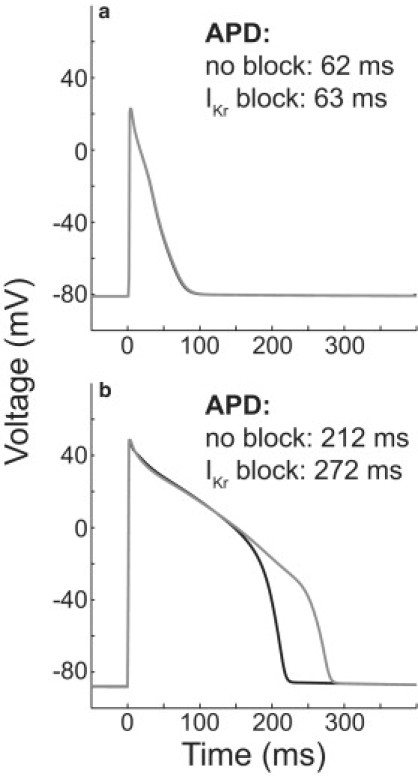
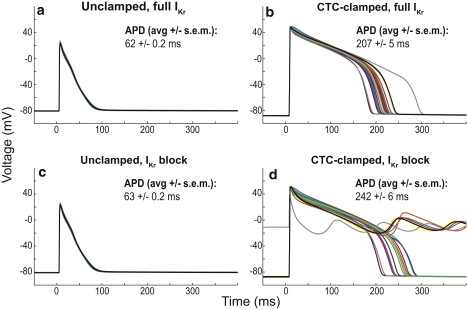
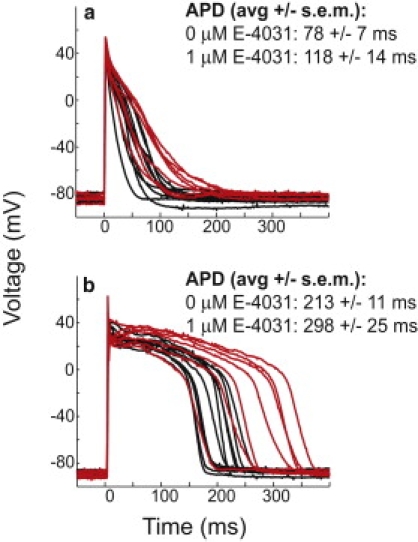
Interspecies differences can limit the translational value of excitable cells isolated from model organisms. It can be difficult to extrapolate from a drug- or mutation-induced phenotype in mice to human pathophysiology because mouse and human cardiac electrodynamics differ greatly. We present a hybrid computational-experimental technique, the cell-type transforming clamp, which is designed to overcome such differences by using a calculated compensatory current to convert the macroscopic electrical behavior of an isolated cell into that of a different cell type. We demonstrate the technique's utility by evaluating drug arrhythmogenicity in murine cardiomyocytes that are transformed to behave like human myocytes. Whereas we use the cell-type transforming clamp in this work to convert between mouse and human electrodynamics, the technique could be adapted to convert between the action potential morphologies of any two cell types of interest.
Figures
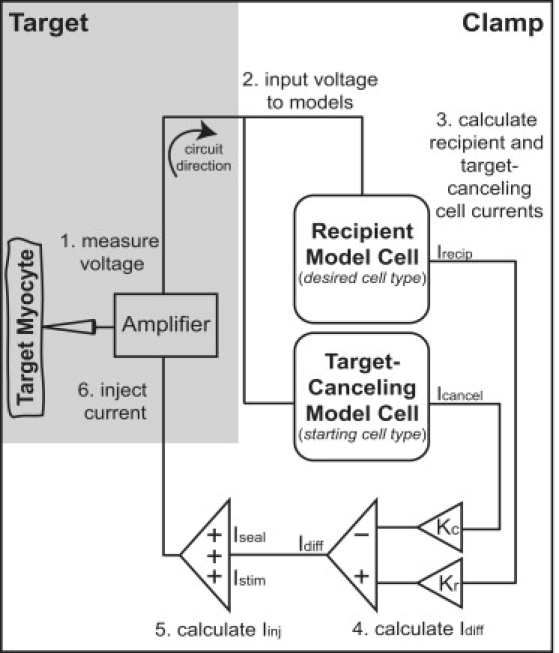
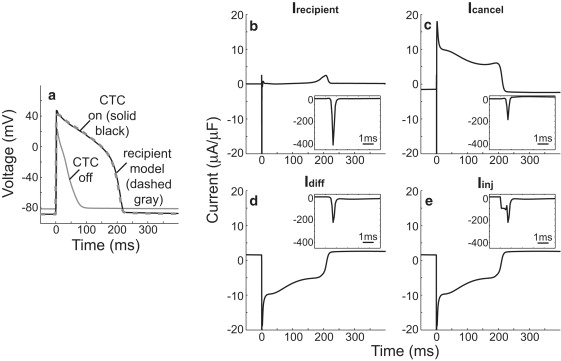
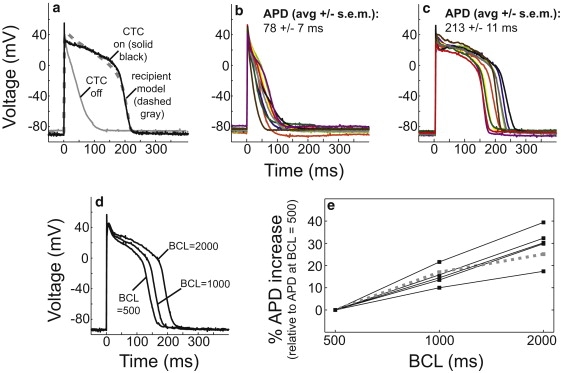
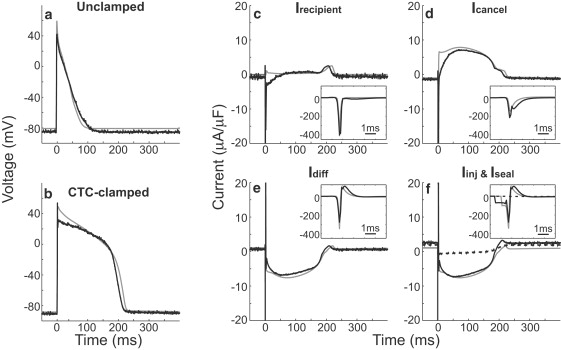
References
-
- Nerbonne J.M. Studying cardiac arrhythmias in the mouse—a reasonable model for probing mechanisms? Trends Cardiovasc. Med. 2004;14:83–93. - PubMed
-
- Doerr T., Denger R., Trautwein W. Calcium currents in single SA nodal cells of the rabbit heart studied with action potential clamp. Pflugers Arch. 1989;413:599–603. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
