

Mechanosensitive closed-closed transitions in large membrane proteins: osmoprotection and tension damping
- PMID: 19917230
- PMCID: PMC2776245
- DOI: 10.1016/j.bpj.2009.08.054
Mechanosensitive closed-closed transitions in large membrane proteins: osmoprotection and tension damping
Abstract
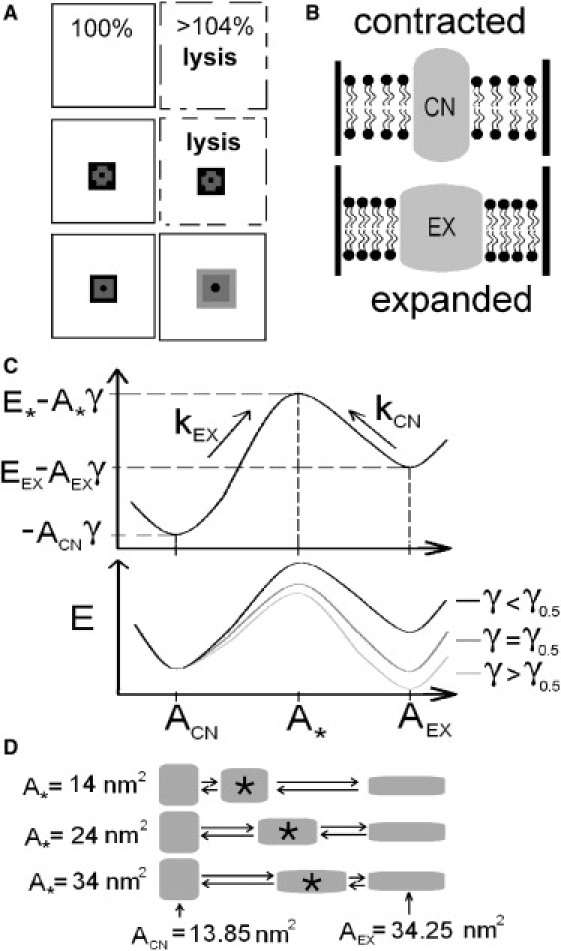
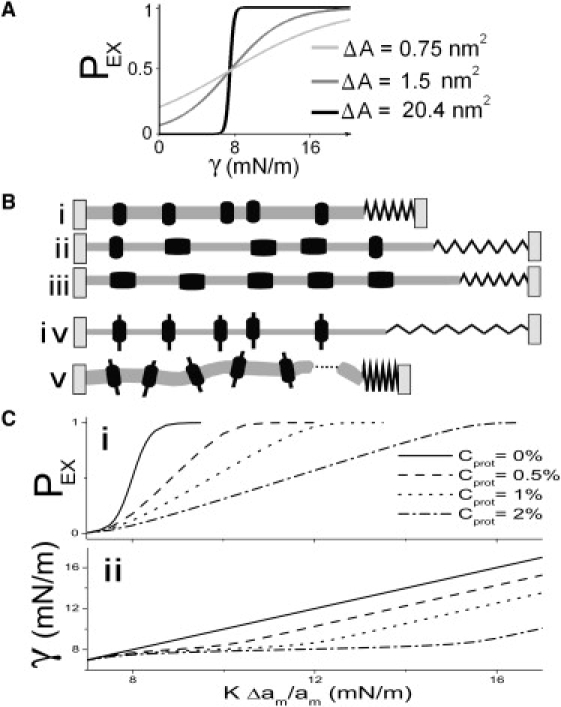
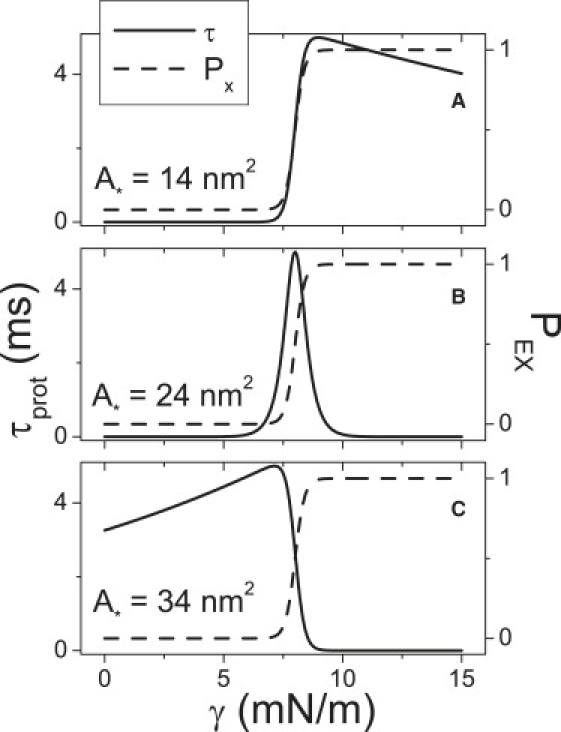
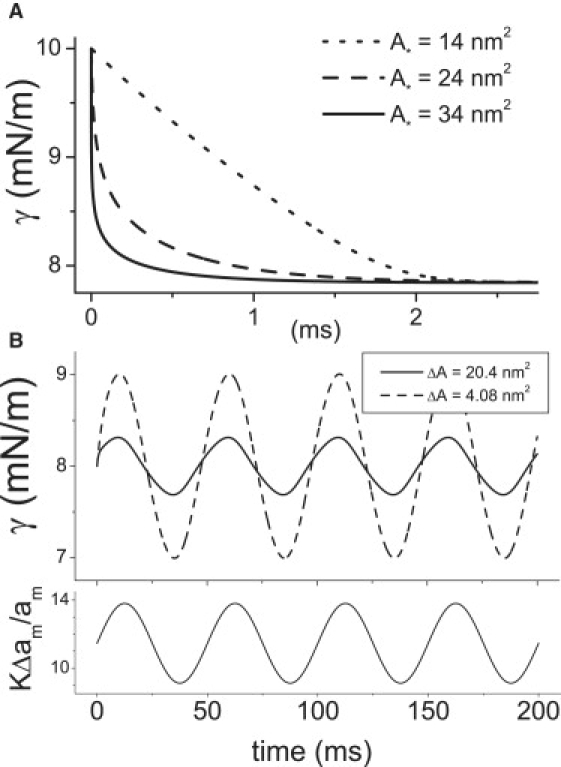
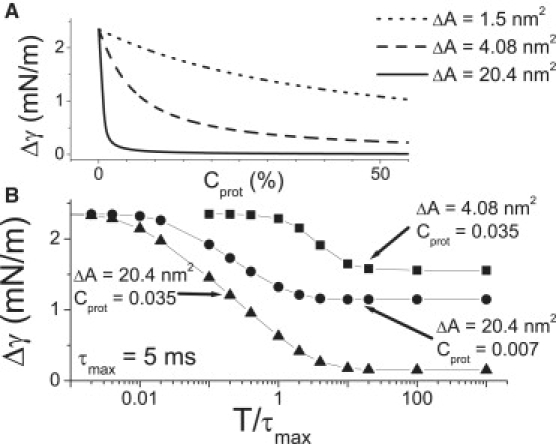
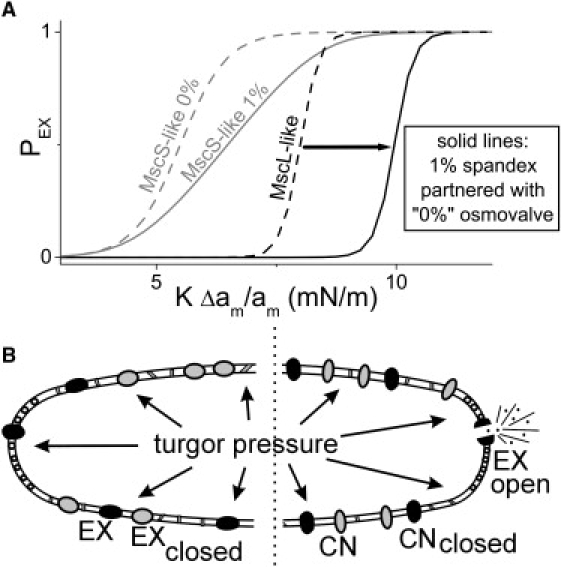
Multiconformation membrane proteins are mechanosensitive (MS) if their conformations displace different bilayer areas. Might MS closed-closed transitions serve as tension buffers, that is, as membrane "spandex"? While bilayer expansion is effectively instantaneous, transitions of bilayer-embedded MS proteins are stochastic (thermally activated) so spandex kinetics would be critical. Here we model generic two-state (contracted/expanded) stochastic spandexes inspired by known bacterial osmovalves (MscL, MscS) then suggest experimental approaches to test for spandex-like behaviors in these proteins. Modeling shows: 1), spandex kinetics depend on the transition state location along an area reaction coordinate; 2), increasing membrane concentration of a spandex right-shifts its midpoint (= tension-Boltzmann); 3), spandexes with midpoints below the activating tension of an osmovalve could optimize osmovalve deployment (required: large midpoint, barrier near the expanded state); 4), spandexes could damp bilayer tension excursions (required: midpoint at target tension, and for speed, barrier halfway between the contracted and expanded states; the larger the spandex Delta-area, the more precise the maintenance of target tension; higher spandex concentrations damp larger amplitude strain fluctuations). One spandex species could not excel as both first line of defense for osmovalve partners and tension damper. Possible interactions among MS closed-closed and closed-open transitions are discussed for MscS- and MscL-like proteins.
Figures
References
-
- Hamill O.P. Twenty odd years of stretch-sensitive channels. Pflügers Arch. Eur. J. Phys. 2006;453:333–351. - PubMed
-
- Sukharev S.I., Anishkin A. Mechanosensitive channels: what can we learn from “simple” model systems? Trends Neurosci. 2004;27:345–351. - PubMed
-
- Booth I.R., Edwards M.D., Black S., Schumann U., Miller S. Mechanosensitive channels in bacteria: signs of closure? Nat. Rev. Microbiol. 2007;5:431–440. - PubMed
-
- Corry B., Martinac B. Bacterial mechanosensitive channels: experiment and theory. Biochim. Biophys. Acta Biomembr. 2008;1778:1859–1870. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
