

Rhythmic PER abundance defines a critical nodal point for negative feedback within the circadian clock mechanism
- PMID: 19917250
- PMCID: PMC3625733
- DOI: 10.1016/j.molcel.2009.10.012
Rhythmic PER abundance defines a critical nodal point for negative feedback within the circadian clock mechanism
Abstract
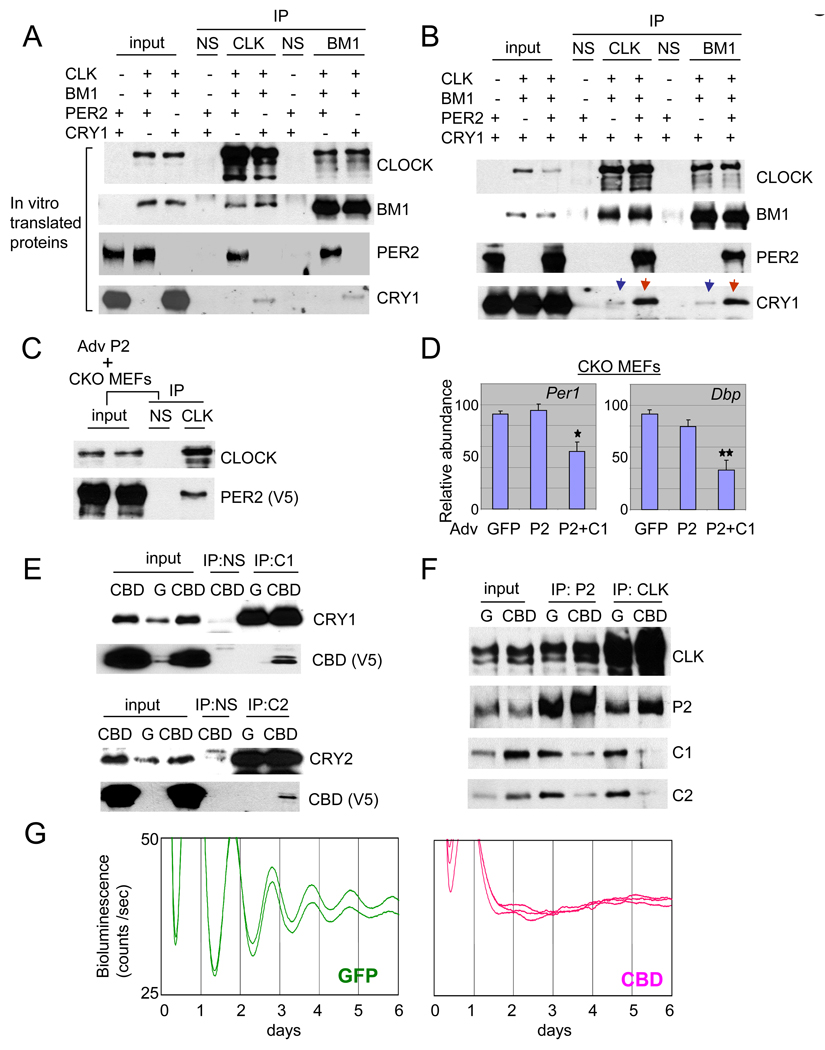
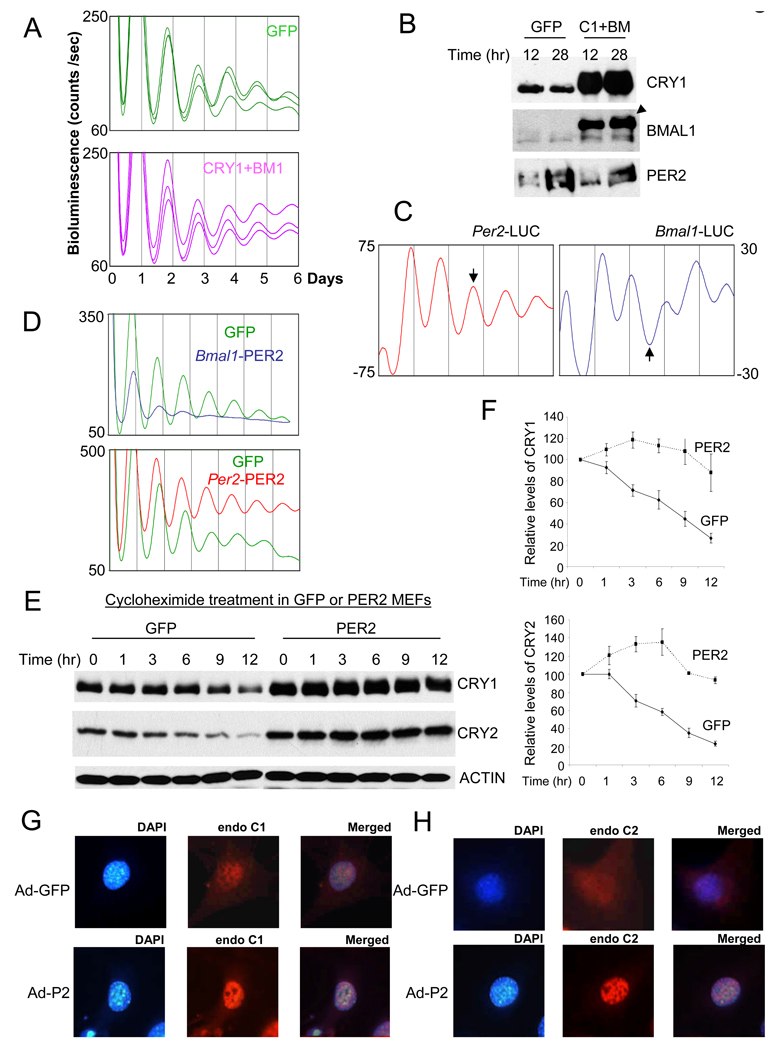
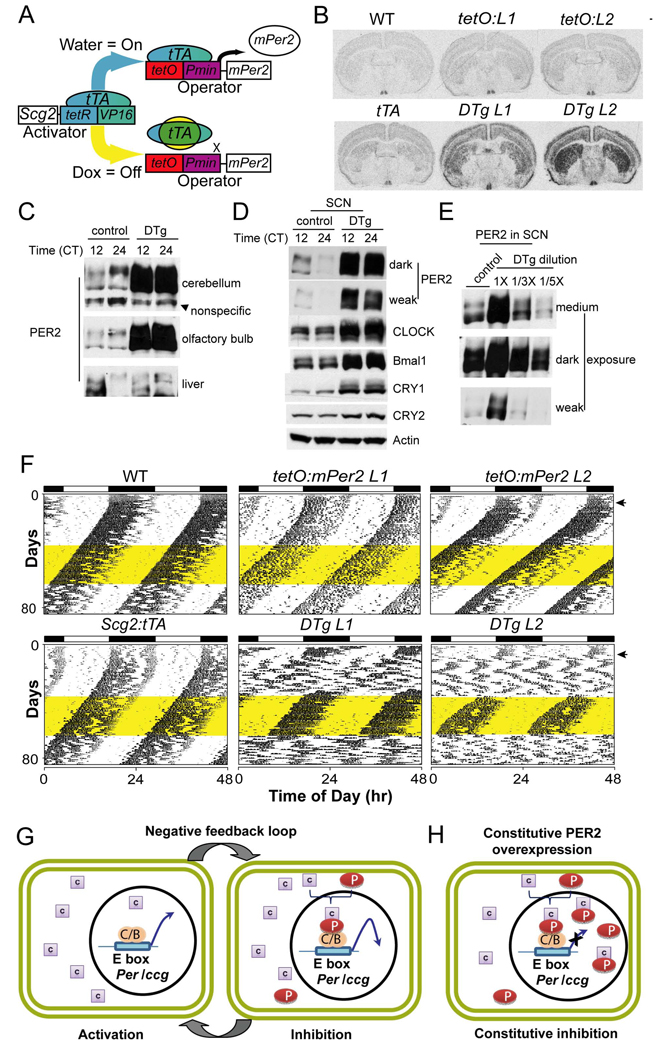
Circadian rhythms in mammals are generated by a transcriptional negative feedback loop that is driven primarily by oscillations of PER and CRY, which inhibit their own transcriptional activators, CLOCK and BMAL1. Current models posit that CRY is the dominant repressor, while PER may play an accessory role. In this study, however, constitutive expression of PER, and not CRY1, severely disrupted the clock in fibroblasts and liver. Furthermore, constitutive expression of PER2 in the brain and SCN of transgenic mice caused a complete loss of behavioral circadian rhythms in a conditional and reversible manner. These results demonstrate that rhythmic levels of PER2, rather than CRY1, are critical for circadian oscillations in cells and in the intact organism. Our biochemical evidence supports an elegant mechanism for the disparity: PER2 directly and rhythmically binds to CLOCK:BMAL1, while CRY only interacts indirectly; PER2 bridges CRY and CLOCK:BMAL1 to drive the circadian negative feedback loop.
Figures
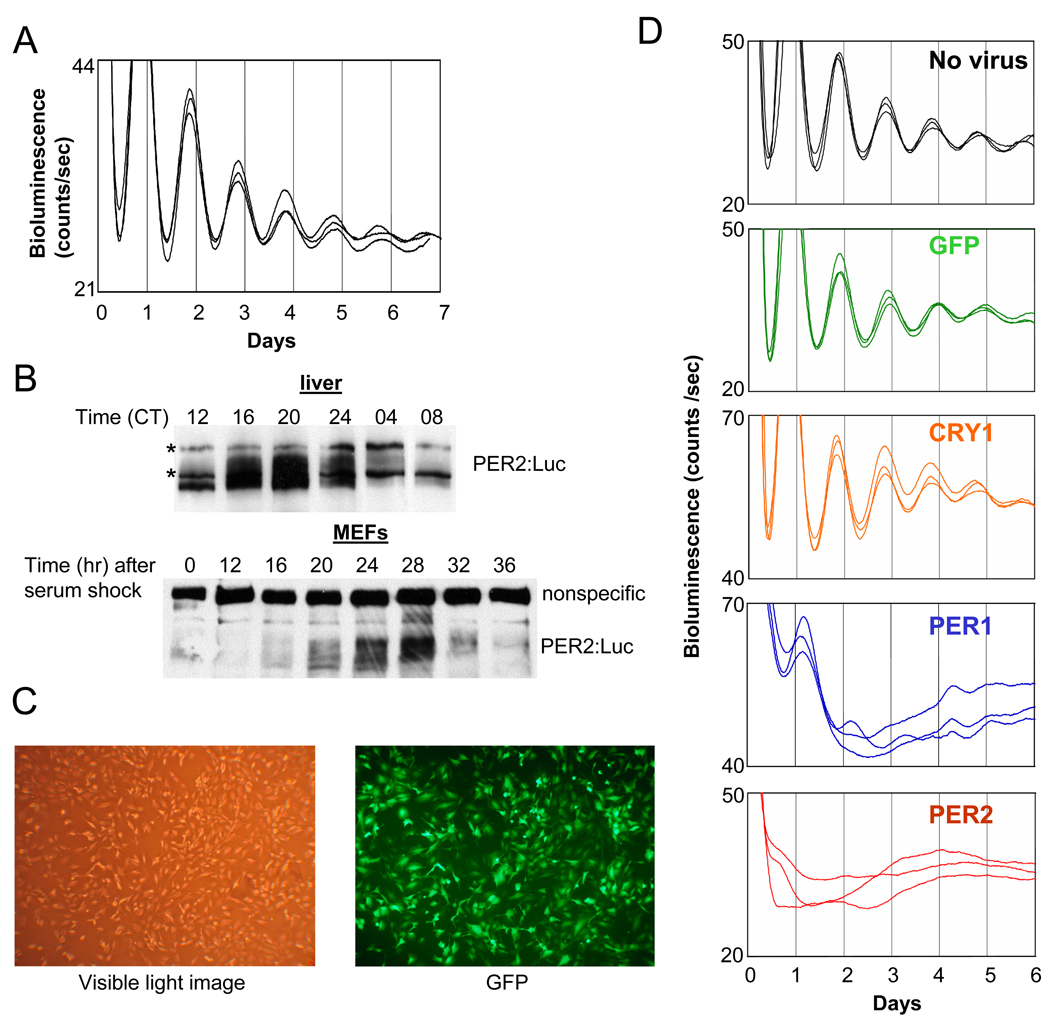
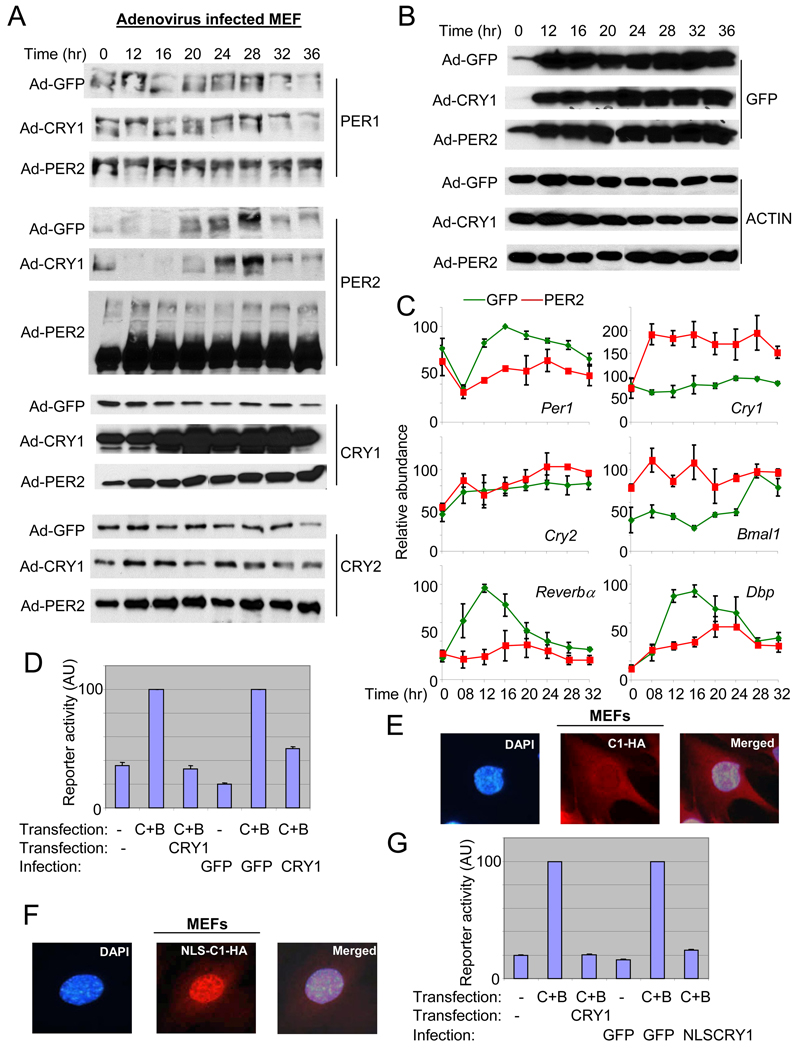
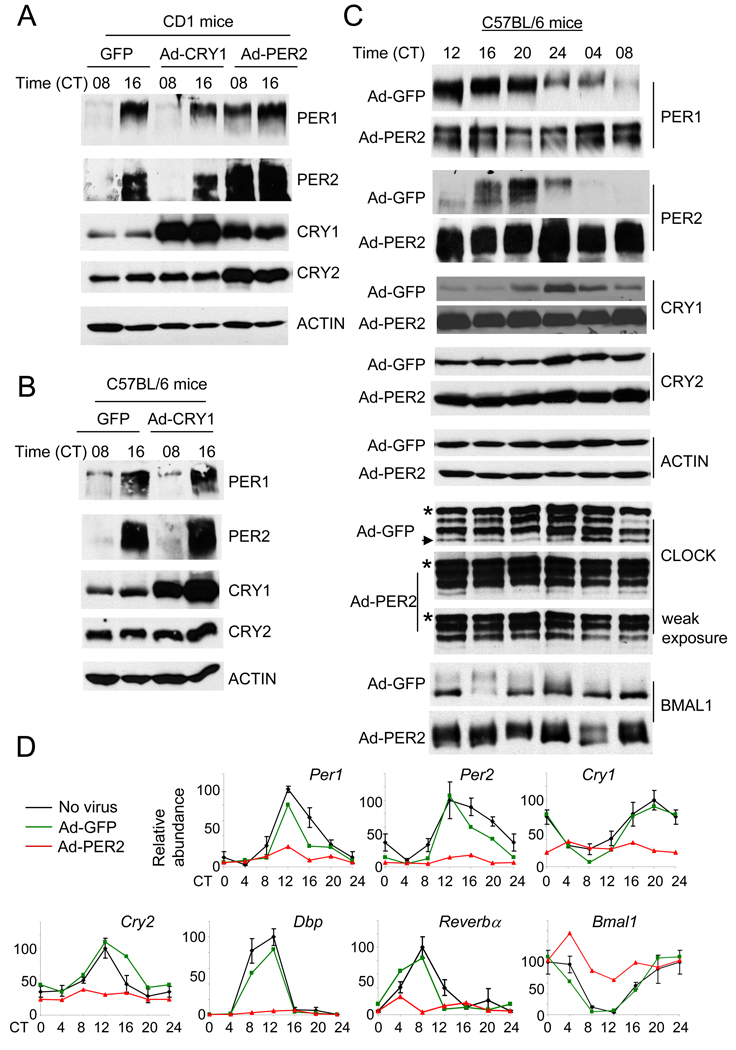
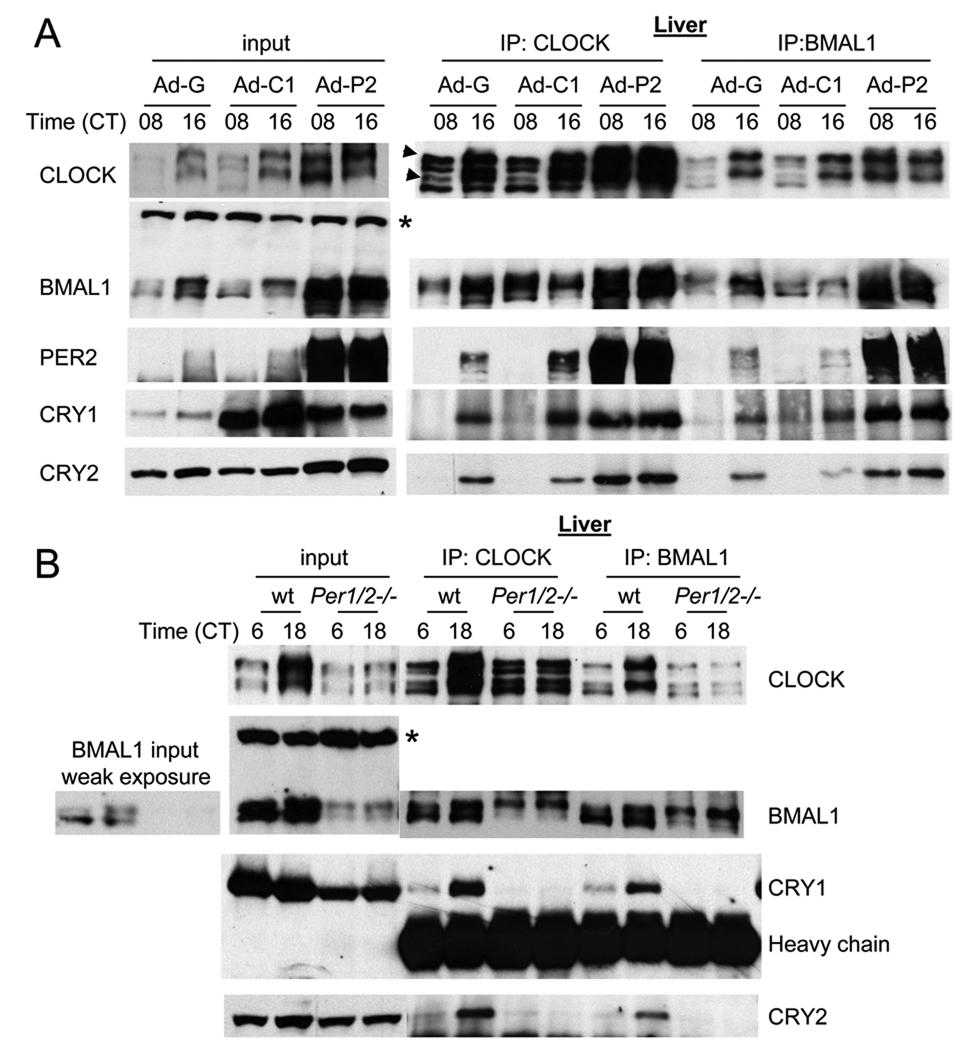
References
-
- Allada R, Emery P, Takahashi JS, Rosbash M. Stopping time: the genetics of fly and mouse circadian clocks. Annu Rev Neurosci. 2001;24:1091–1119. - PubMed
-
- Aronson BD, Johnson KA, Loros JJ, Dunlap JC. Negative feedback defining a circadian clock: autoregulation of the clock gene frequency. Science. 1994;263:1578–1584. - PubMed
-
- Bae K, Jin X, Maywood ES, Hastings MH, Reppert SM, Weaver DR. Differential functions of mPer1, mPer2, and mPer3 in the SCN circadian clock. Neuron. 2001;30:525–536. - PubMed
-
- Baylies MK, Bargiello TA, Jackson FR, Young MW. Changes in abundance or structure of the per gene product can alter periodicity of the Drosophila clock. Nature. 1987;326:390–392. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
