

Visualizing changes in electron distribution in coupled chains of cytochrome bc(1) by modifying barrier for electron transfer between the FeS cluster and heme c(1)
- PMID: 19917265
- PMCID: PMC2807467
- DOI: 10.1016/j.bbabio.2009.11.003
Visualizing changes in electron distribution in coupled chains of cytochrome bc(1) by modifying barrier for electron transfer between the FeS cluster and heme c(1)
Abstract
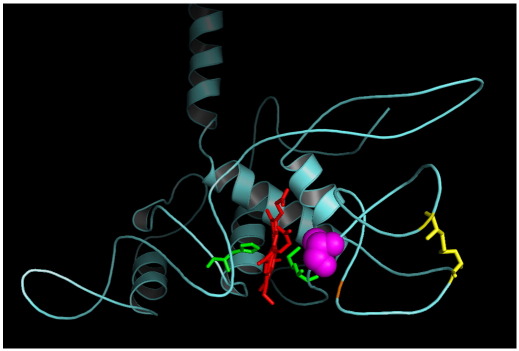
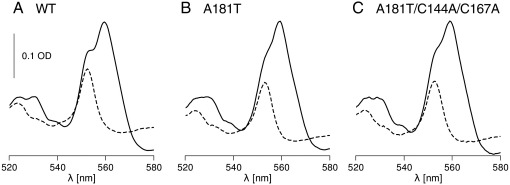
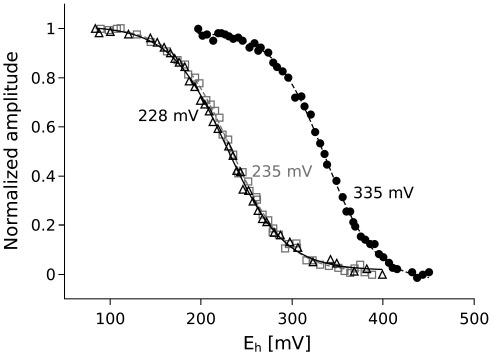
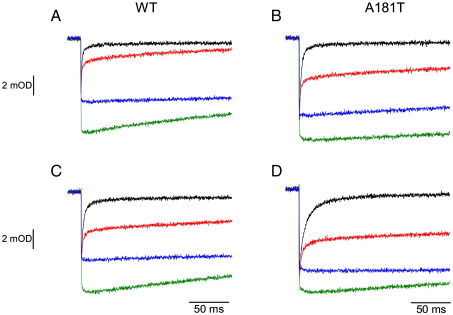
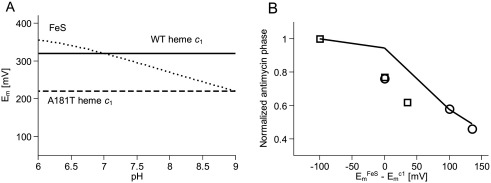
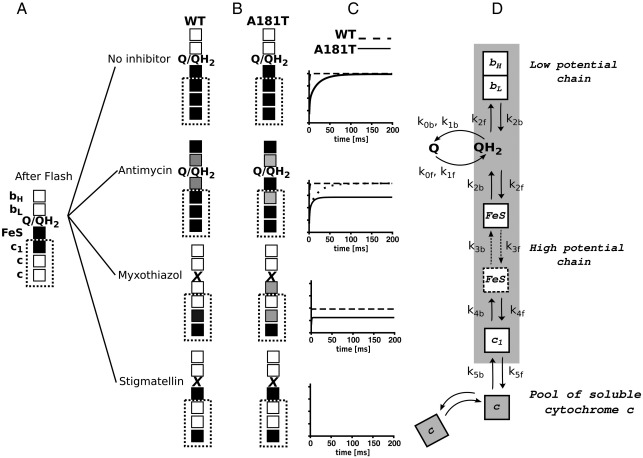
Cytochrome c(1) of Rhodobacter (Rba.) species provides a series of mutants which change barriers for electron transfer through the cofactor chains of cytochrome bc(1) by modifying heme c(1) redox midpoint potential. Analysis of post-flash electron distribution in such systems can provide useful information about the contribution of individual reactions to the overall electron flow. In Rba. capsulatus, the non-functional low-potential forms of cytochrome c(1) which are devoid of the disulfide bond naturally present in this protein revert spontaneously by introducing a second-site suppression (mutation A181T) that brings the potential of heme c(1) back to the functionally high levels, yet maintains it some 100 mV lower from the native value. Here we report that the disulfide and the mutation A181T can coexist in one protein but the mutation exerts a dominant effect on the redox properties of heme c(1) and the potential remains at the same lower value as in the disulfide-free form. This establishes effective means to modify a barrier for electron transfer between the FeS cluster and heme c(1) without breaking disulfide. A comparison of the flash-induced electron transfers in native and mutated cytochrome bc(1) revealed significant differences in the post-flash equilibrium distribution of electrons only when the connection of the chains with the quinone pool was interrupted at the level of either of the catalytic sites by the use of specific inhibitors, antimycin or myxothiazol. In the non-inhibited system no such differences were observed. We explain the results using a kinetic model in which a shift in the equilibrium of one reaction influences the equilibrium of all remaining reactions in the cofactor chains. It follows a rather simple description in which the direction of electron flow through the coupled chains of cytochrome bc(1) exclusively depends on the rates of all reversible partial reactions, including the Q/QH2 exchange rate to/from the catalytic sites.
2009 Elsevier B.V. All rights reserved.
Figures
Similar articles
-
Functional flexibility of electron flow between quinol oxidation Qo site of cytochrome bc1 and cytochrome c revealed by combinatory effects of mutations in cytochrome b, iron-sulfur protein and cytochrome c1.Biochim Biophys Acta Bioenerg. 2018 Sep;1859(9):754-761. doi: 10.1016/j.bbabio.2018.04.010. Epub 2018 Apr 27. Biochim Biophys Acta Bioenerg. 2018. PMID: 29705394
-
Electron sweep across four b-hemes of cytochrome bc1 revealed by unusual paramagnetic properties of the Qi semiquinone intermediate.Biochim Biophys Acta Bioenerg. 2018 Jun;1859(6):459-469. doi: 10.1016/j.bbabio.2018.03.010. Epub 2018 Mar 27. Biochim Biophys Acta Bioenerg. 2018. PMID: 29596789
-
Magnetic interactions sense changes in distance between heme b(L) and the iron-sulfur cluster in cytochrome bc(1).Biochemistry. 2009 Jun 23;48(24):5708-20. doi: 10.1021/bi900511b. Biochemistry. 2009. PMID: 19415898 Free PMC article.
-
Role of the Rieske iron-sulfur protein midpoint potential in the protonmotive Q-cycle mechanism of the cytochrome bc1 complex.J Bioenerg Biomembr. 1999 Jun;31(3):235-42. doi: 10.1023/a:1005419712731. J Bioenerg Biomembr. 1999. PMID: 10591529 Review.
-
Inter- and intra-molecular electron transfer in the cytochrome bc(1) complex.Biochim Biophys Acta. 2002 Sep 10;1555(1-3):65-70. doi: 10.1016/s0005-2728(02)00256-6. Biochim Biophys Acta. 2002. PMID: 12206893 Review.
Cited by
-
Tyrosine triad at the interface between the Rieske iron-sulfur protein, cytochrome c1 and cytochrome c2 in the bc1 complex of Rhodobacter capsulatus.Biochim Biophys Acta. 2012 May;1817(5):811-8. doi: 10.1016/j.bbabio.2012.01.013. Epub 2012 Jan 28. Biochim Biophys Acta. 2012. PMID: 22306765 Free PMC article.
-
The High-Spin Heme b L Mutant Exposes Dominant Reaction Leading to the Formation of the Semiquinone Spin-Coupled to the [2Fe-2S]+ Cluster at the Qo Site of Rhodobacter capsulatus Cytochrome bc 1.Front Chem. 2021 May 7;9:658877. doi: 10.3389/fchem.2021.658877. eCollection 2021. Front Chem. 2021. PMID: 34026724 Free PMC article.
-
Mitochondrial disease-related mutations at the cytochrome b-iron-sulfur protein (ISP) interface: Molecular effects on the large-scale motion of ISP and superoxide generation studied in Rhodobacter capsulatus cytochrome bc1.Biochim Biophys Acta. 2016 Aug;1857(8):1102-1110. doi: 10.1016/j.bbabio.2016.03.022. Epub 2016 Mar 28. Biochim Biophys Acta. 2016. PMID: 27032290 Free PMC article.
-
Binding of imidazole to the heme of cytochrome c1 and inhibition of the bc1 complex from Rhodobacter sphaeroides: II. Kinetics and mechanism of binding.J Biol Chem. 2010 Jul 16;285(29):22522-31. doi: 10.1074/jbc.M110.128082. Epub 2010 May 6. J Biol Chem. 2010. PMID: 20448037 Free PMC article.
-
Discrimination between two possible reaction sequences that create potential risk of generation of deleterious radicals by cytochrome bc₁. Implications for the mechanism of superoxide production.Biochim Biophys Acta. 2010 Nov;1797(11):1820-7. doi: 10.1016/j.bbabio.2010.07.005. Epub 2010 Jul 15. Biochim Biophys Acta. 2010. PMID: 20637719 Free PMC article.
References
-
- Mitchell P. Protonmotive redox mechanism of the cytochrome b-c1 complex in the respiratory chain: protonmotive ubiquinone cycle. FEBS Lett. 1975;56:1–6. - PubMed
-
- Brandt U., Trumpower B.L. The protonmotive Q cycle in mitochondria and bacteria. Crit. Rev. Biochem. Mol. Biol. 1994;29:165–197. - PubMed
-
- Berry E.A., Guergova-Kuras M., Huang L., Crofts A.R. Structure and function of cytochrome bc complexes. Annu. Rev. Biochem. 2000;69:1005–1075. - PubMed
-
- Cape J.L., Bowman M.K., Kramer D.M. Understanding the cytochrome bc complexes by what they don't do. The Q cycle at 30. Trends Plant Sci. 2006;11:46–55. - PubMed
-
- Crofts A.R. Proton pumping in the bc1 complex: a new gating mechanism that prevents short circuits. Biochim. Biophys. Acta. 2006;1757:1019–1034. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
