

An electrostatic interaction between TEA and an introduced pore aromatic drives spring-in-the-door inactivation in Shaker potassium channels
- PMID: 19917730
- PMCID: PMC2806421
- DOI: 10.1085/jgp.200910260
An electrostatic interaction between TEA and an introduced pore aromatic drives spring-in-the-door inactivation in Shaker potassium channels
Abstract
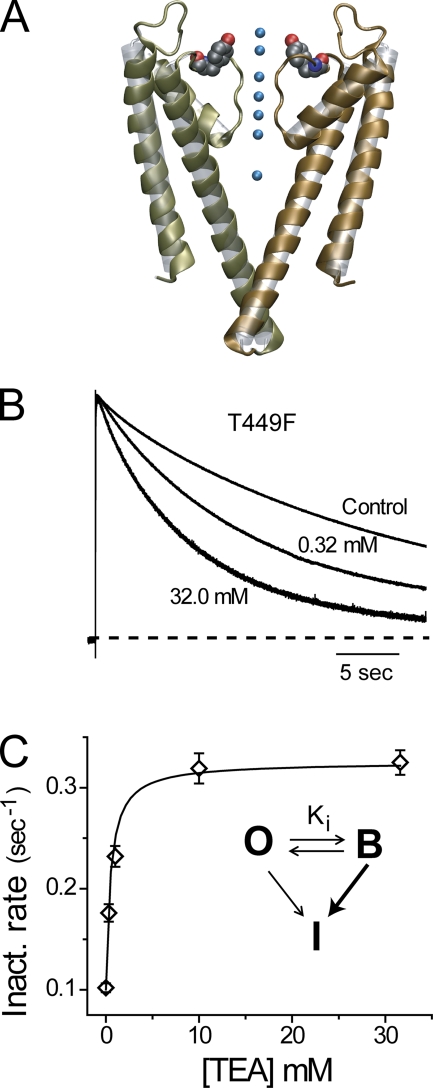
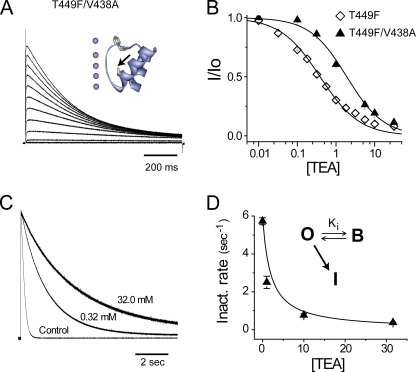
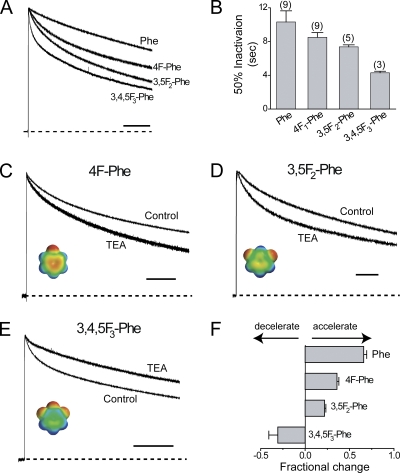
Slow inactivation of Kv1 channels involves conformational changes near the selectivity filter. We examine such changes in Shaker channels lacking fast inactivation by considering the consequences of mutating two residues, T449 just external to the selectivity filter and V438 in the pore helix near the bottom of the selectivity filter. Single mutant T449F channels with the native V438 inactivate very slowly, and the canonical foot-in-the-door effect of extracellular tetraethylammonium (TEA) is not only absent, but the time course of slow inactivation is accelerated by TEA. The V438A mutation dramatically speeds inactivation in T449F channels, and TEA slows inactivation exactly as predicted by the foot-in-the-door model. We propose that TEA has this effect on V438A/T449F channels because the V438A mutation produces allosteric consequences within the selectivity filter and may reorient the aromatic ring at position 449. We investigated the possibility that the blocker promotes the collapse of the outer vestibule (spring-in-the-door) in single mutant T449F channels by an electrostatic attraction between a cationic TEA and the quadrupole moments of the four aromatic rings. To test this idea, we used in vivo nonsense suppression to serially fluorinate the introduced aromatic ring at the 449 position, a manipulation that withdraws electrons from the aromatic face with little effect on the shape, net charge, or hydrophobicity of the aromatic ring. Progressive fluorination causes monotonically enhanced rates of inactivation. In further agreement with our working hypothesis, increasing fluorination of the aromatic gradually transforms the TEA effect from spring-in-the-door to foot-in-the-door. We further substantiate our electrostatic hypothesis by quantum mechanical calculations.
Figures

Comment in
-
It's spring-time for slow inactivation.J Gen Physiol. 2009 Dec;134(6):457-9. doi: 10.1085/jgp.200910353. Epub 2009 Nov 16. J Gen Physiol. 2009. PMID: 19917731 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
