

Structure of deviations from optimality in biological systems
- PMID: 19918070
- PMCID: PMC2777958
- DOI: 10.1073/pnas.0905336106
Structure of deviations from optimality in biological systems
Abstract
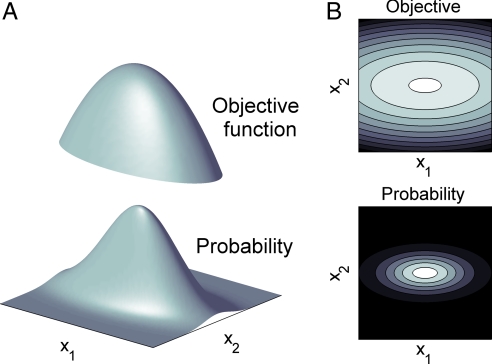
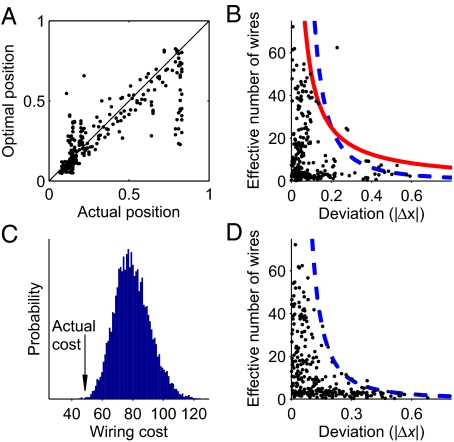
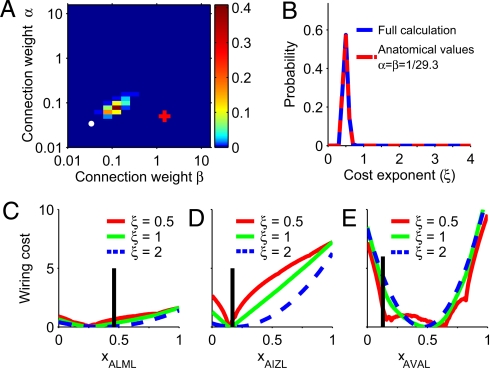
Optimization theory has been used to analyze evolutionary adaptation. This theory has explained many features of biological systems, from the genetic code to animal behavior. However, these systems show important deviations from optimality. Typically, these deviations are large in some particular components of the system, whereas others seem to be almost optimal. Deviations from optimality may be due to many factors in evolution, including stochastic effects and finite time, that may not allow the system to reach the ideal optimum. However, we still expect the system to have a higher probability of reaching a state with a higher value of the proposed indirect measure of fitness. In systems of many components, this implies that the largest deviations are expected in those components with less impact on the indirect measure of fitness. Here, we show that this simple probabilistic rule explains deviations from optimality in two very different biological systems. In Caenorhabditis elegans, this rule successfully explains the experimental deviations of the position of neurons from the configuration of minimal wiring cost. In Escherichia coli, the probabilistic rule correctly obtains the structure of the experimental deviations of metabolic fluxes from the configuration that maximizes biomass production. This approach is proposed to explain or predict more data than optimization theory while using no extra parameters. Thus, it can also be used to find and refine hypotheses about which constraints have shaped biological structures in evolution.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Barton NH, Briggs DEG, Eisen JA, Goldstein DB, Patel NH. Evolution. Cold Spring Harbor, NY: Cold Spring Harbor Lab Press; 2007.
-
- Alexander RM. Optima for Animals. Princeton: Princeton Univ Press; 1996.
-
- Parker GA, Maynard Smith J. Optimality theory in evolutionary biology. Nature. 1990;348:27–33.
-
- Freeland SJ, Hurst LD. The genetic code is one in a million. J Mol Evol. 1998;47:238–248. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
