

A theory of the influence of eye movements on the refinement of direction selectivity in the cat's primary visual cortex
- PMID: 19919281
- PMCID: PMC2951336
- DOI: 10.3109/09548980903314204
A theory of the influence of eye movements on the refinement of direction selectivity in the cat's primary visual cortex
Abstract
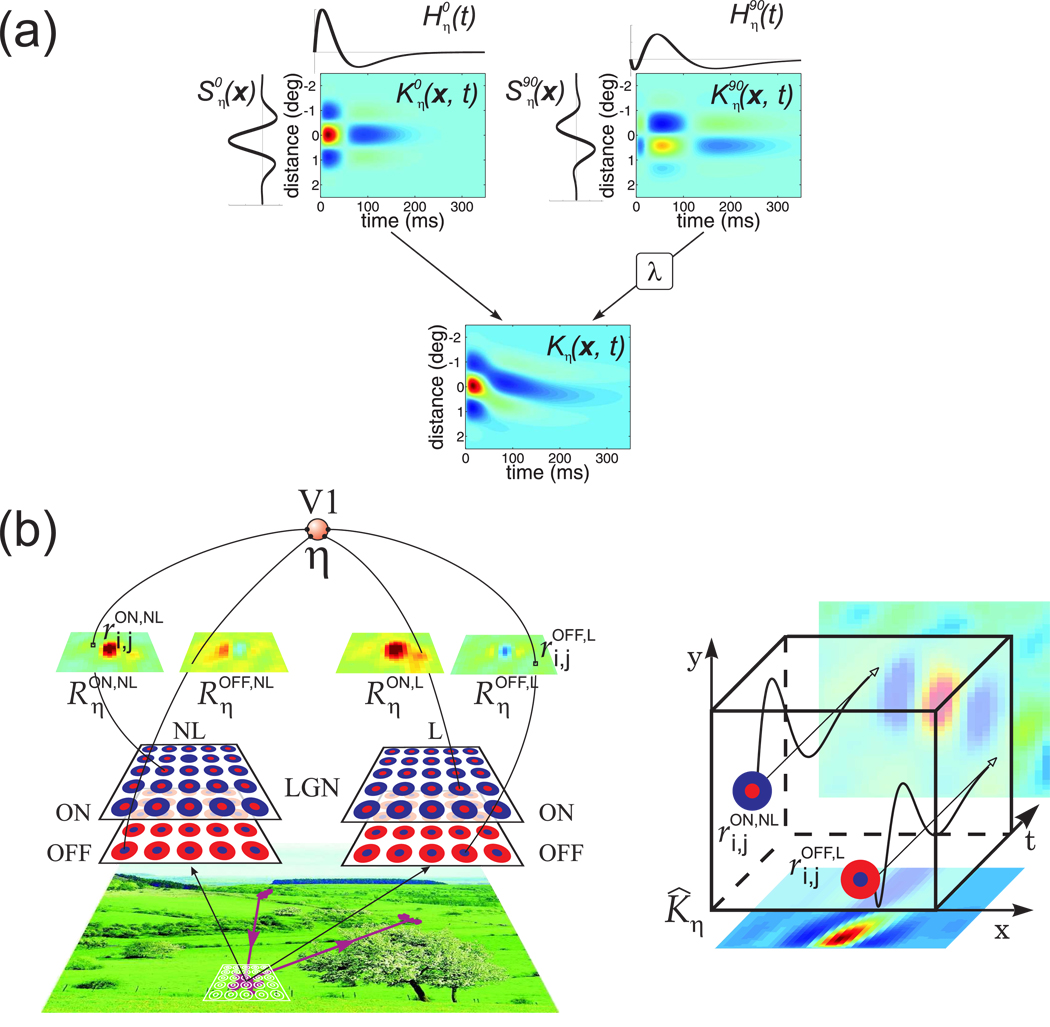
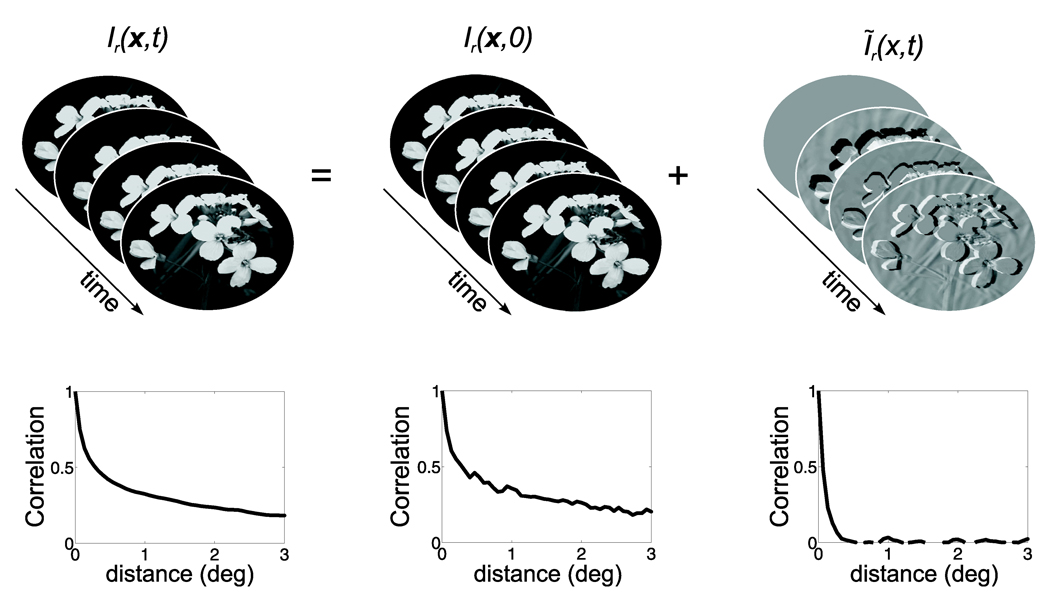
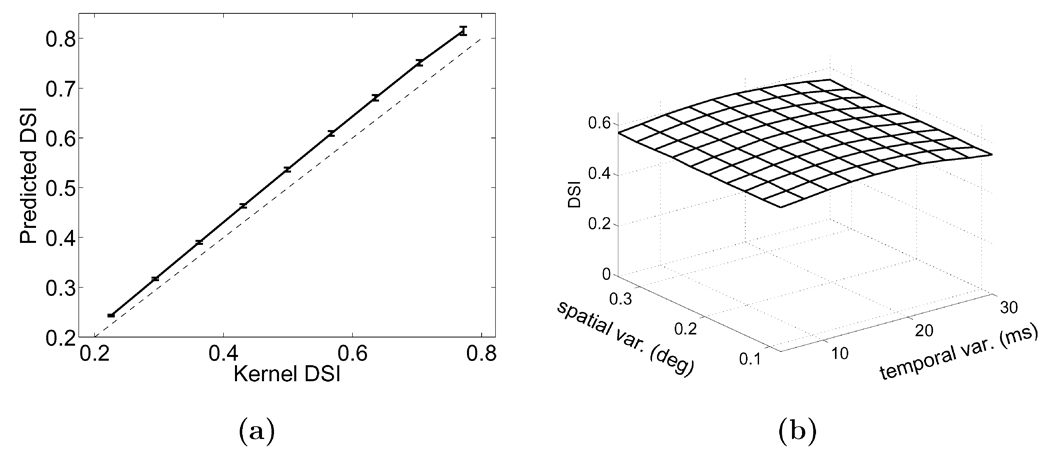
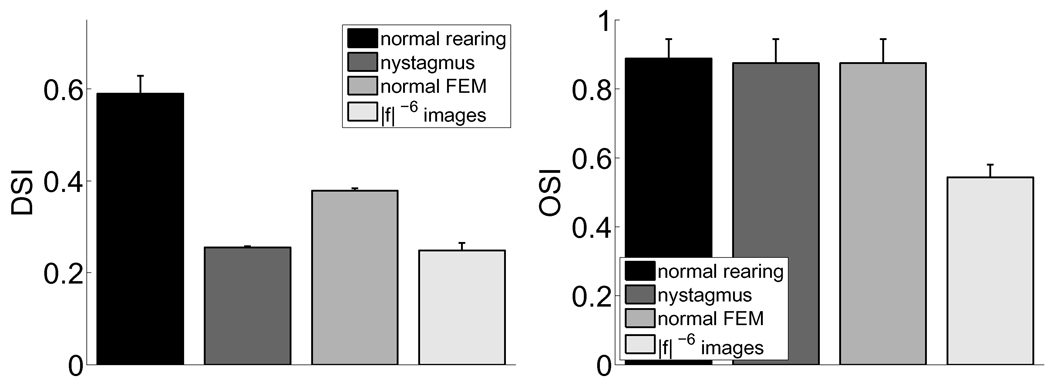
Early in life, visual experience influences the refinement of the preferential response for specific stimulus features exhibited by neurons in the primary visual cortex. A striking example of this influence is the reduction in cortical direction selectivity observed in cats reared under high-frequency stroboscopic illumination. Although various mechanisms have been proposed to explain the maturation of individual properties of neuronal responses, a unified account of the joint development of the multiple response features of cortical neurons has remained elusive. In this study, we show that Hebbian synaptic plasticity accounts for the simultaneous refinement of orientation and direction selectivity under both normal and stroboscopic rearing, if one takes into account the spatiotemporal input to the retina during oculomotor activity. In a computational model of the LGN and V1, eye movements are sufficient to establish the patterns of thalamocortical activity required for a Hebbian refinement of both direction- and orientation-selective responses during exposure to natural stimuli. Furthermore, we show that consideration of fixational eye movements explains the simultaneous loss of direction selectivity and preservation of orientation selectivity observed as a consequence of stroboscopic rearing. These results further support a role for oculomotor activity in the refinement of the response properties of V1 neurons.
Figures





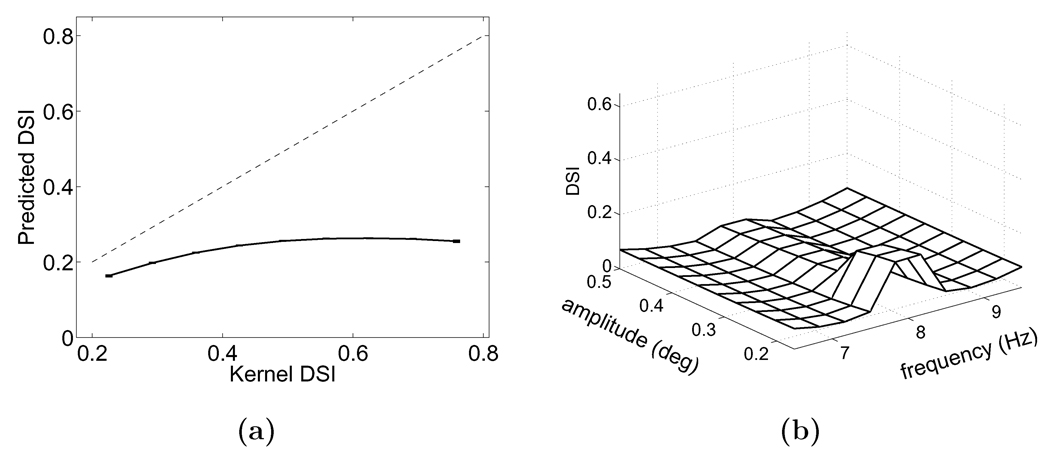
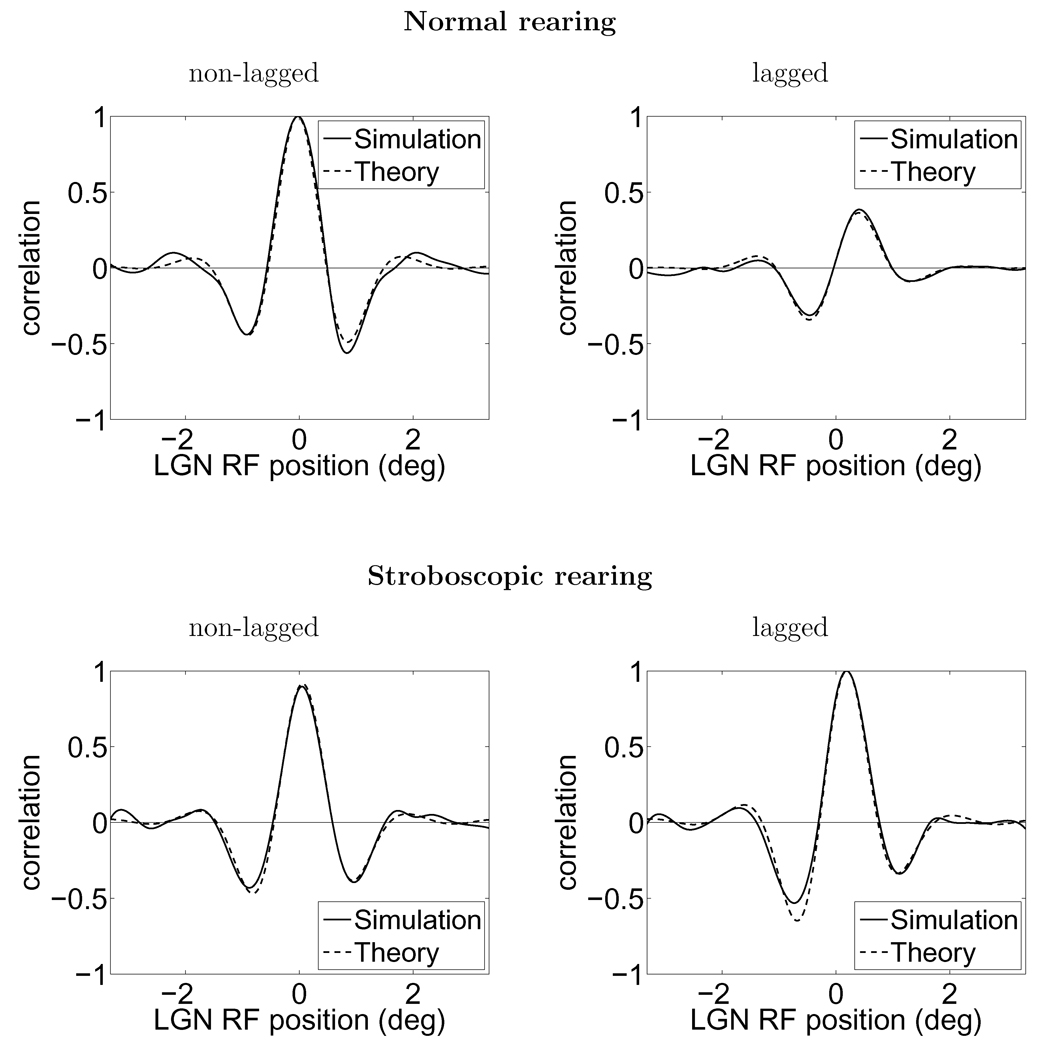

Similar articles
-
Decorrelation of neural activity during fixational instability: possible implications for the refinement of V1 receptive fields.Vis Neurosci. 2004 Sep-Oct;21(5):725-38. doi: 10.1017/S0952523804215073. Vis Neurosci. 2004. PMID: 15683560
-
A theoretical analysis of the influence of fixational instability on the development of thalamocortical connectivity.Neural Comput. 2006 Mar;18(3):569-90. doi: 10.1162/089976606775623270. Neural Comput. 2006. PMID: 16483408
-
Orientation tuning of surround suppression in lateral geniculate nucleus and primary visual cortex of cat.Neuroscience. 2007 Nov 23;149(4):962-75. doi: 10.1016/j.neuroscience.2007.08.001. Epub 2007 Aug 9. Neuroscience. 2007. PMID: 17945429
-
The dynamics of visual responses in the primary visual cortex.Prog Brain Res. 2007;165:21-32. doi: 10.1016/S0079-6123(06)65003-6. Prog Brain Res. 2007. PMID: 17925238 Review.
-
Development of neuronal selectivity in primary visual cortex of cat.Physiol Rev. 1984 Jan;64(1):325-434. doi: 10.1152/physrev.1984.64.1.325. Physiol Rev. 1984. PMID: 6320235 Review. No abstract available.
Cited by
-
Contrast sensitivity reveals an oculomotor strategy for temporally encoding space.Elife. 2019 Jan 8;8:e40924. doi: 10.7554/eLife.40924. Elife. 2019. PMID: 30620333 Free PMC article.
-
The unsteady eye: an information-processing stage, not a bug.Trends Neurosci. 2015 Apr;38(4):195-206. doi: 10.1016/j.tins.2015.01.005. Epub 2015 Feb 16. Trends Neurosci. 2015. PMID: 25698649 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous