

Substrate specificity of three cytochrome c haem lyase isoenzymes from Wolinella succinogenes: unconventional haem c binding motifs are not sufficient for haem c attachment by NrfI and CcsA1
- PMID: 19919672
- PMCID: PMC3414424
- DOI: 10.1111/j.1365-2958.2009.06965.x
Substrate specificity of three cytochrome c haem lyase isoenzymes from Wolinella succinogenes: unconventional haem c binding motifs are not sufficient for haem c attachment by NrfI and CcsA1
Abstract
Bacterial c-type cytochrome maturation is dependent on a complex enzymic machinery. The key reaction is catalysed by cytochrome c haem lyase (CCHL) that usually forms two thioether bonds to attach haem b to the cysteine residues of a haem c binding motif (HBM) which is, in most cases, a CX(2)CH sequence. Here, the HBM specificity of three distinct CCHL isoenzymes (NrfI, CcsA1 and CcsA2) from the Epsilonproteobacterium Wolinella succinogenes was investigated using either W. succinogenes or Escherichia coli as host organism. Several reporter c-type cytochromes were employed including cytochrome c nitrite reductases (NrfA) from E. coli and Campylobacter jejuni that differ in their active-site HBMs (CX(2)CK or CX(2)CH). W. succinogenes CcsA2 was found to attach haem to standard CX(2)CH motifs in various cytochromes whereas other HBMs were not recognized. NrfI was able to attach haem c to the active-site CX(2)CK motif of both W. succinogenes and E. coli NrfA, but not to NrfA from C. jejuni. Different apo-cytochrome variants carrying the CX(15)CH motif, assumed to be recognized by CcsA1 during maturation of the octahaem cytochrome MccA, were not processed by CcsA1 in either W. succinogenes or E. coli. It is concluded that the dedicated CCHLs NrfI and CcsA1 attach haem to non-standard HBMs only in the presence of further, as yet uncharacterized structural features. Interestingly, it proved impossible to delete the ccsA2 gene from the W. succinogenes genome, a finding that is discussed in the light of the available genomic, proteomic and functional data on W. succinogenes c-type cytochromes.
Figures
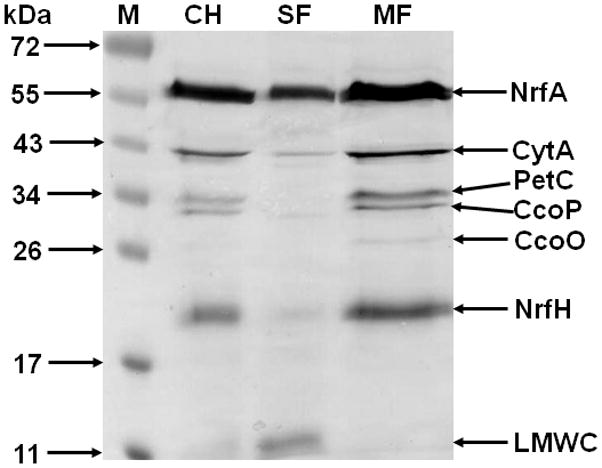
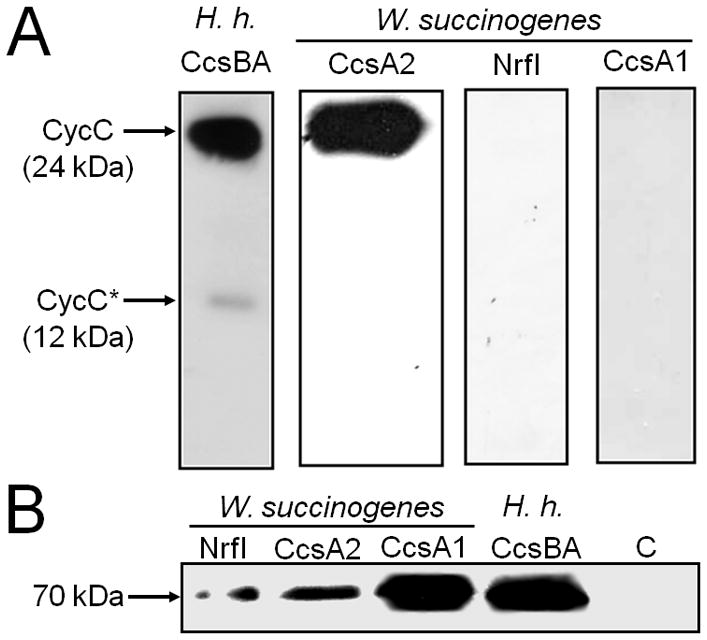
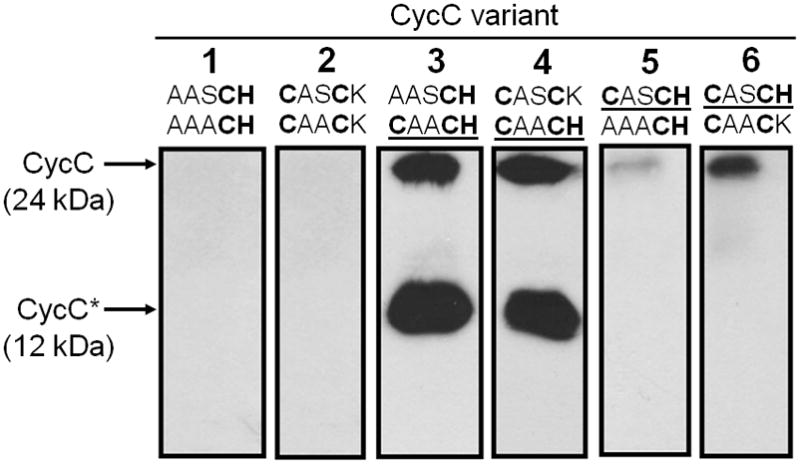
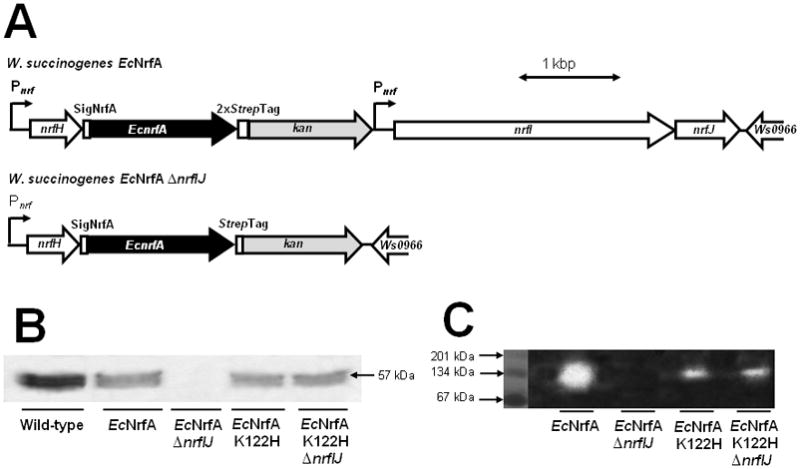
References
-
- Ahuja U, Kjelgaard P, Schulz BL, Thöny-Meyer L, Hederstedt L. Haem-delivery proteins in cytochrome c maturation system II. Mol Microbiol. 2009;73:1058–1071. - PubMed
-
- Allen JWA, Sawyer EB, Ginger ML, Barker PD, Ferguson SJ. Variant c-type cytochromes as probes of the substrate specificity of the E. coli cytochrome c maturation (Ccm) apparatus. Biochem J. 2009;419:177–184. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
