

Therapeutic potential of adult bone marrow-derived mesenchymal stem cells in prostate cancer bone metastasis
- PMID: 19920103
- PMCID: PMC2943933
- DOI: 10.1158/1078-0432.CCR-09-1938
Therapeutic potential of adult bone marrow-derived mesenchymal stem cells in prostate cancer bone metastasis
Abstract
Purpose: Current evidence indicates that an osteoblast lesion in prostate cancer is preceded by osteolysis. Thus, prevention of osteolysis would reduce complications of bone metastasis. Bone marrow-derived mesenchymal stem cells have the ability to differentiate into osteoblast and produce osteoprotegerin, a decoy receptor for the receptor activator for nuclear factor kappaB ligand, naturally. The present study examined the potential of unmodified mesenchymal stem cells to prevent osteolytic bone lesions in a preclinical mouse model of prostate cancer.
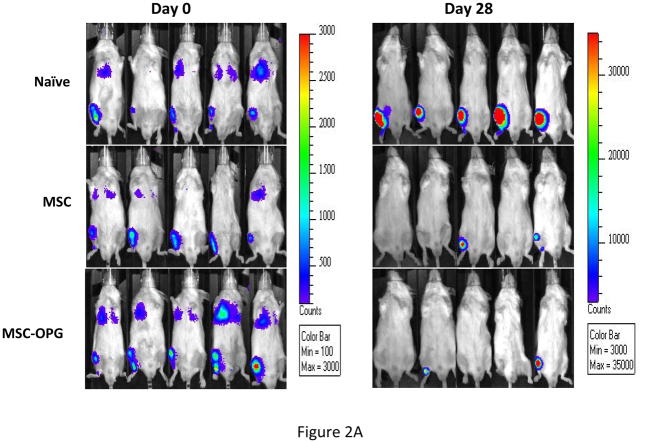
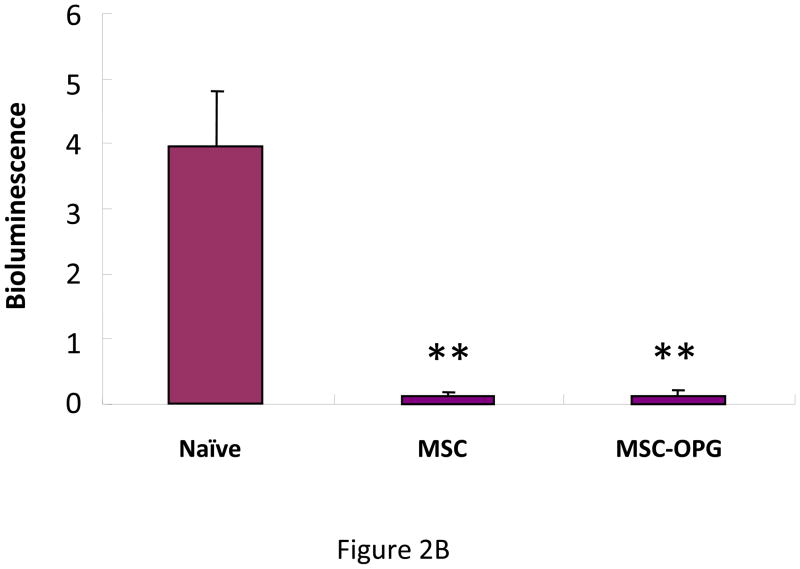
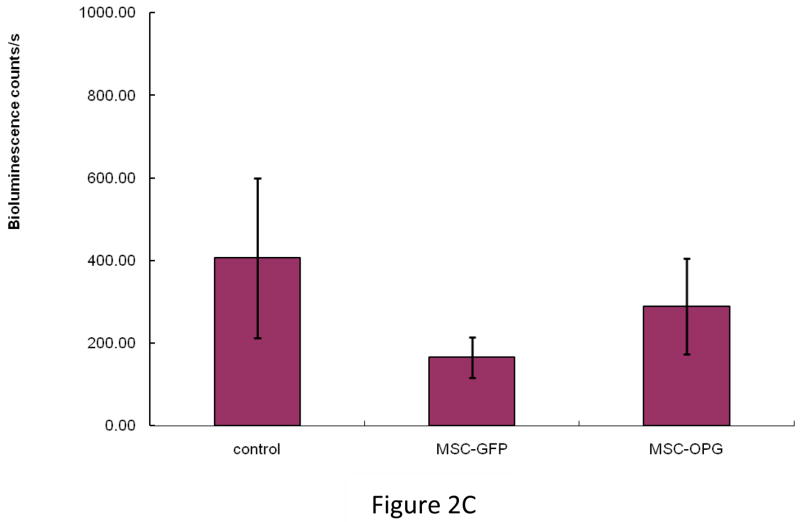
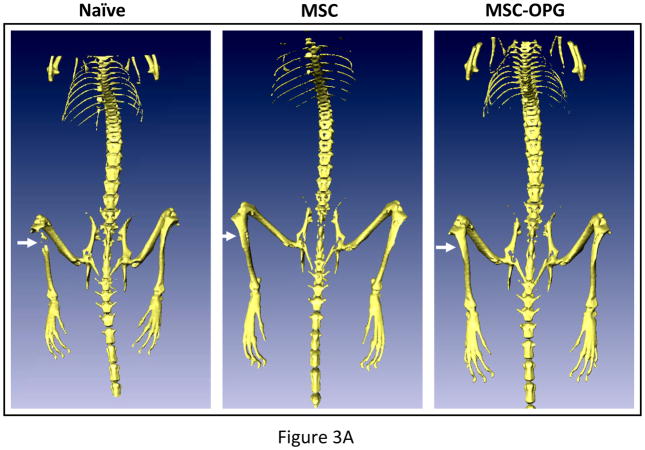
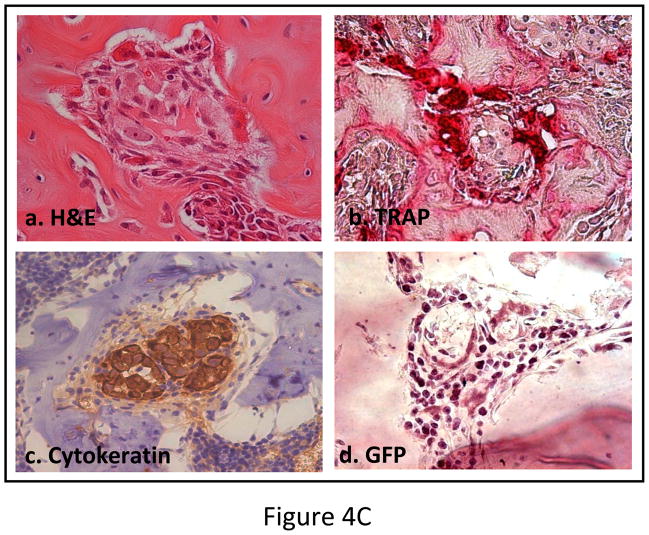
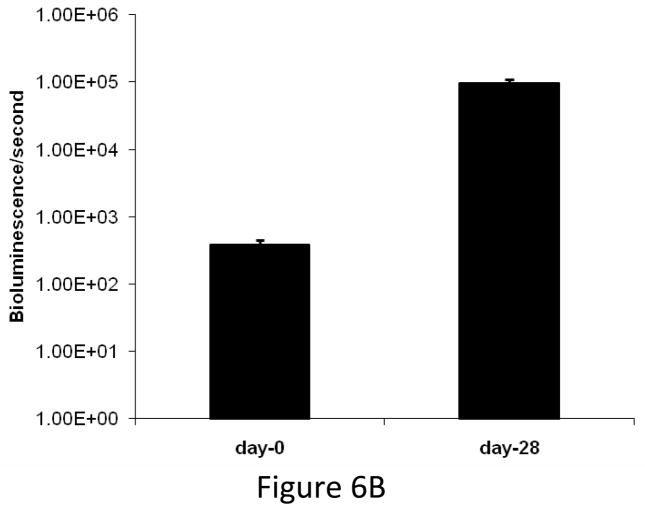
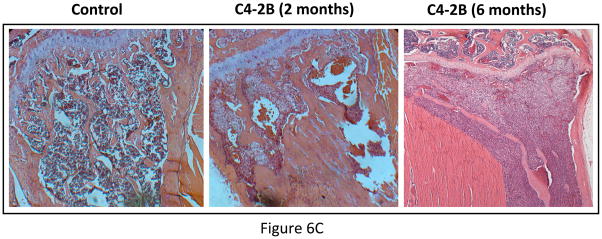
Experimental design: The human prostate cancer cell line PC3 was implanted in tibiae of severe combined immunodeficient mice. After establishment of the tumor, either unmodified or genetically engineered mesenchymal stem cells overexpressing osteoprotegerin was injected at the site of tumor growth. The effects of therapy were monitored by bioluminescence imaging, micro-computed tomography, immunohistochemistry, and histomorphometry.
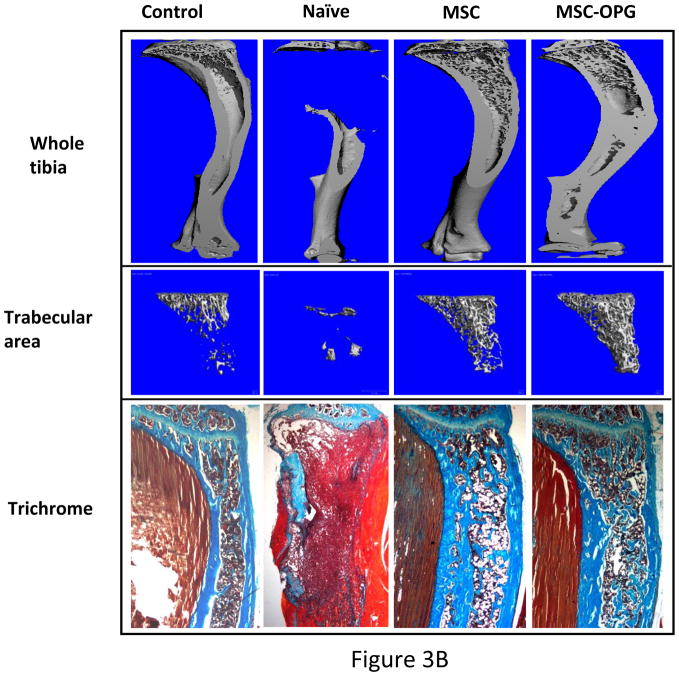
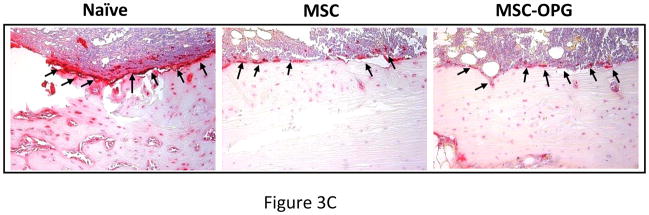
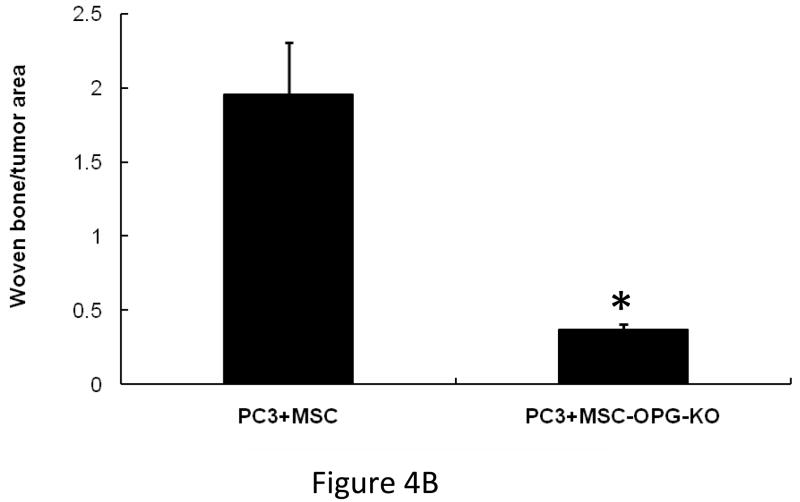
Results: Data indicated significant (P < 0.001) inhibition of tumor growth and restoration of bone in mice treated with unmodified and modified mesenchymal stem cells. Detailed analysis suggested that the donor mesenchymal stem cell inhibited tumor progression by producing woven bone around the growing tumor cells in the tibiae and by preventing osteoclastogenesis.
Conclusions: Overcoming the limitation of the number of mesenchymal stem cells available in the bone can provide significant amelioration for osteolytic damage without further modification.
Figures

Similar articles
-
Severe combined immunodeficient-hu model of human prostate cancer metastasis to human bone.Cancer Res. 1999 Apr 15;59(8):1987-93. Cancer Res. 1999. PMID: 10213511
-
Antitumoral activity and osteogenic potential of mesenchymal stem cells expressing the urokinase-type plasminogen antagonist amino-terminal fragment in a murine model of osteolytic tumor.Stem Cells. 2008 Nov;26(11):2981-90. doi: 10.1634/stemcells.2008-0139. Epub 2008 Aug 28. Stem Cells. 2008. PMID: 18757301
-
Combined inhibition of the BMP pathway and the RANK-RANKL axis in a mixed lytic/blastic prostate cancer lesion.Bone. 2011 Mar 1;48(3):578-87. doi: 10.1016/j.bone.2010.11.003. Epub 2010 Nov 10. Bone. 2011. PMID: 21073986 Free PMC article.
-
The role of osteoclastic activity in prostate cancer skeletal metastases.Drugs Today (Barc). 2002 Feb;38(2):91-102. doi: 10.1358/dot.2002.38.2.820105. Drugs Today (Barc). 2002. PMID: 12532187 Review.
-
RANK-Fc: a therapeutic antagonist for RANK-L in myeloma.Cancer. 2003 Feb 1;97(3 Suppl):802-12. doi: 10.1002/cncr.11134. Cancer. 2003. PMID: 12548579 Review.
Cited by
-
Prostate cancer and bone: the elective affinities.Biomed Res Int. 2014;2014:167035. doi: 10.1155/2014/167035. Epub 2014 May 28. Biomed Res Int. 2014. PMID: 24971315 Free PMC article. Review.
-
Depletion of plasmacytoid dendritic cells inhibits tumor growth and prevents bone metastasis of breast cancer cells.J Immunol. 2012 Nov 1;189(9):4258-65. doi: 10.4049/jimmunol.1101855. Epub 2012 Sep 26. J Immunol. 2012. PMID: 23018462 Free PMC article.
-
Human mesenchymal stem cells inhibit metastasis of a hepatocellular carcinoma model using the MHCC97-H cell line.Cancer Sci. 2010 Dec;101(12):2546-53. doi: 10.1111/j.1349-7006.2010.01738.x. Epub 2010 Oct 7. Cancer Sci. 2010. PMID: 20942864 Free PMC article.
-
Inhibition of Human Malignant Pleural Mesothelioma Growth by Mesenchymal Stromal Cells.Cells. 2021 Jun 8;10(6):1427. doi: 10.3390/cells10061427. Cells. 2021. PMID: 34201002 Free PMC article.
-
Inhibition of uPAR-TGFβ crosstalk blocks MSC-dependent EMT in melanoma cells.J Mol Med (Berl). 2015 Jul;93(7):783-94. doi: 10.1007/s00109-015-1266-2. Epub 2015 Feb 20. J Mol Med (Berl). 2015. PMID: 25694039
References
-
- Jemal A, Siegel R, Ward E, Murray T, Xu H, Thun MJ. Cancer statistics. CA Cancer J Clin. 2007;57:43–66. - PubMed
-
- Ye L, Kynaston HG, Jiang WG. Bone metastasis in prostate cancer: molecular and cellular mechanisms. Int J Mol Med. 2007;20:103–11. - PubMed
-
- Guise TA, Yin JJ, Mohammad KS. Role of endothelin-1 in osteoblastic bone metastases. Cancer. 2003;97:779–84. - PubMed
-
- Choueiri MB, Tu SM, Yu-Lee LY, Lin SH. The central role of osteoblasts in the metastasis of prostate cancer. Cancer Metastasis Rev. 2006;25:601–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
