

Yeast cohesin complex embraces 2 micron plasmid sisters in a tri-linked catenane complex
- PMID: 19920123
- PMCID: PMC2811031
- DOI: 10.1093/nar/gkp993
Yeast cohesin complex embraces 2 micron plasmid sisters in a tri-linked catenane complex
Abstract
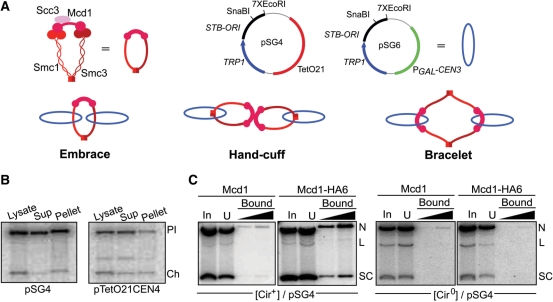
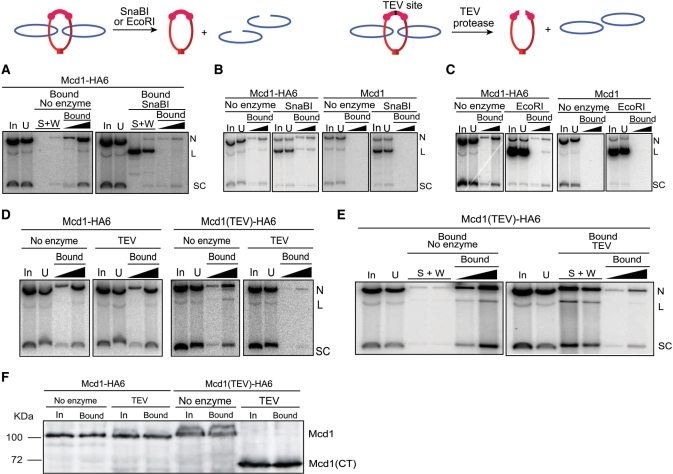
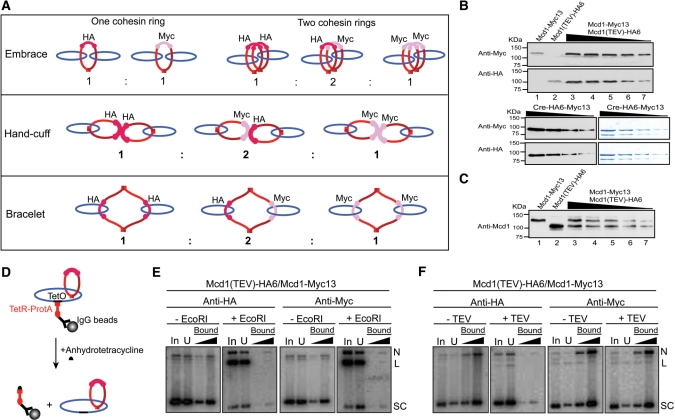
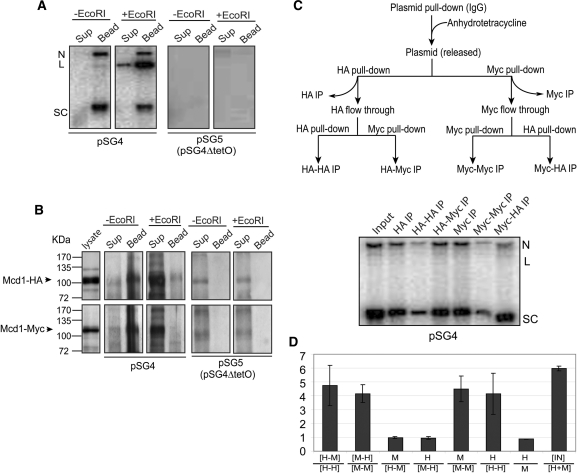
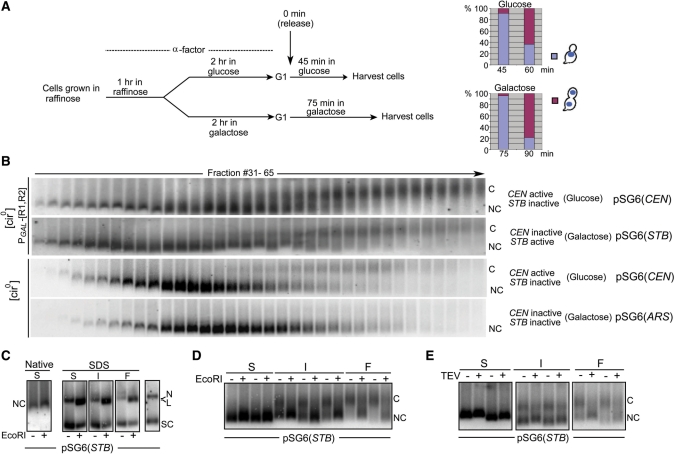
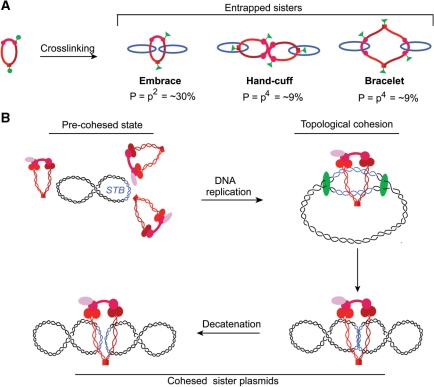
Sister chromatid cohesion, crucial for faithful segregation of replicated chromosomes in eukaryotes, is mediated by the multi-subunit protein complex cohesin. The Saccharomyces cerevisiae plasmid 2 micron circle mimics chromosomes in assembling cohesin at its partitioning locus. The plasmid is a multi-copy selfish DNA element that resides in the nucleus and propagates itself stably, presumably with assistance from cohesin. In metaphase cell lysates, or fractions enriched for their cohesed state by sedimentation, plasmid molecules are trapped topologically by the protein ring formed by cohesin. They can be released from cohesin's embrace either by linearizing the DNA or by cleaving a cohesin subunit. Assays using two distinctly tagged cohesin molecules argue against the hand-cuff (an associated pair of monomeric cohesin rings) or the bracelet (a dimeric cohesin ring) model as responsible for establishing plasmid cohesion. Our cumulative results most easily fit a model in which a single monomeric cohesin ring, rather than a series of such rings, conjoins a pair of sister plasmids. These features of plasmid cohesion account for its sister-to-sister mode of segregation by cohesin disassembly during anaphase. The mechanistic similarities of cohesion between mini-chromosome sisters and 2 micron plasmid sisters suggest a potential kinship between the plasmid partitioning locus and centromeres.
Figures
Similar articles
-
The cohesin ring concatenates sister DNA molecules.Nature. 2008 Jul 17;454(7202):297-301. doi: 10.1038/nature07098. Epub 2008 Jul 2. Nature. 2008. PMID: 18596691
-
Mutations in a partitioning protein and altered chromatin structure at the partitioning locus prevent cohesin recruitment by the Saccharomyces cerevisiae plasmid and cause plasmid missegregation.Mol Cell Biol. 2004 Jun;24(12):5290-303. doi: 10.1128/MCB.24.12.5290-5303.2004. Mol Cell Biol. 2004. PMID: 15169893 Free PMC article.
-
Cohesion is established during DNA replication utilising chromosome associated cohesin rings as well as those loaded de novo onto nascent DNAs.Elife. 2020 Jun 9;9:e56611. doi: 10.7554/eLife.56611. Elife. 2020. PMID: 32515737 Free PMC article.
-
The torments of the cohesin ring.Nucleus. 2017 May 4;8(3):261-267. doi: 10.1080/19491034.2017.1295200. Epub 2017 Feb 27. Nucleus. 2017. PMID: 28453390 Free PMC article. Review.
-
Chromosome cohesion - rings, knots, orcs and fellowship.J Cell Sci. 2008 Jul 1;121(Pt 13):2107-14. doi: 10.1242/jcs.029132. J Cell Sci. 2008. PMID: 18565823 Review.
Cited by
-
A selfish DNA element engages a meiosis-specific motor and telomeres for germ-line propagation.J Cell Biol. 2014 Jun 9;205(5):643-61. doi: 10.1083/jcb.201312002. J Cell Biol. 2014. PMID: 24914236 Free PMC article.
-
Histone H3-variant Cse4-induced positive DNA supercoiling in the yeast plasmid has implications for a plasmid origin of a chromosome centromere.Proc Natl Acad Sci U S A. 2011 Aug 16;108(33):13671-6. doi: 10.1073/pnas.1101944108. Epub 2011 Aug 1. Proc Natl Acad Sci U S A. 2011. PMID: 21807992 Free PMC article.
-
Centromere identity: a challenge to be faced.Mol Genet Genomics. 2010 Aug;284(2):75-94. doi: 10.1007/s00438-010-0553-4. Epub 2010 Jun 29. Mol Genet Genomics. 2010. PMID: 20585957 Review.
-
The yeast 2-micron plasmid Rep2 protein has Rep1-independent partitioning function.Nucleic Acids Res. 2022 Oct 14;50(18):10571-10585. doi: 10.1093/nar/gkac810. Nucleic Acids Res. 2022. PMID: 36156142 Free PMC article.
-
Cse4 (CenH3) association with the Saccharomyces cerevisiae plasmid partitioning locus in its native and chromosomally integrated states: implications in centromere evolution.Mol Cell Biol. 2011 Mar;31(5):1030-40. doi: 10.1128/MCB.01191-10. Epub 2010 Dec 20. Mol Cell Biol. 2011. PMID: 21173161 Free PMC article.
References
-
- Onn I, Heidinger-Pauli JM, Guacci V, Unal E, Koshland DE. Sister chromatid cohesion: a simple concept with a complex reality. Annu. Rev. Cell Dev. Biol. 2008;24:105–129. - PubMed
-
- Uhlmann F. The mechanism of sister chromatid cohesion. Exp. Cell Res. 2004;296:80–85. - PubMed
-
- Watanabe Y. Sister chromatid cohesion along arms and at centromeres. Trends Genet. 2005;21:405–412. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
