

The FGFRL1 receptor is shed from cell membranes, binds fibroblast growth factors (FGFs), and antagonizes FGF signaling in Xenopus embryos
- PMID: 19920134
- PMCID: PMC2804375
- DOI: 10.1074/jbc.M109.058248
The FGFRL1 receptor is shed from cell membranes, binds fibroblast growth factors (FGFs), and antagonizes FGF signaling in Xenopus embryos
Abstract
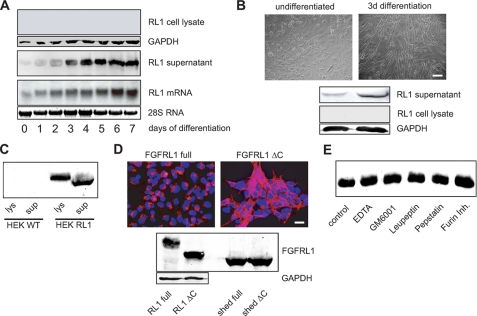
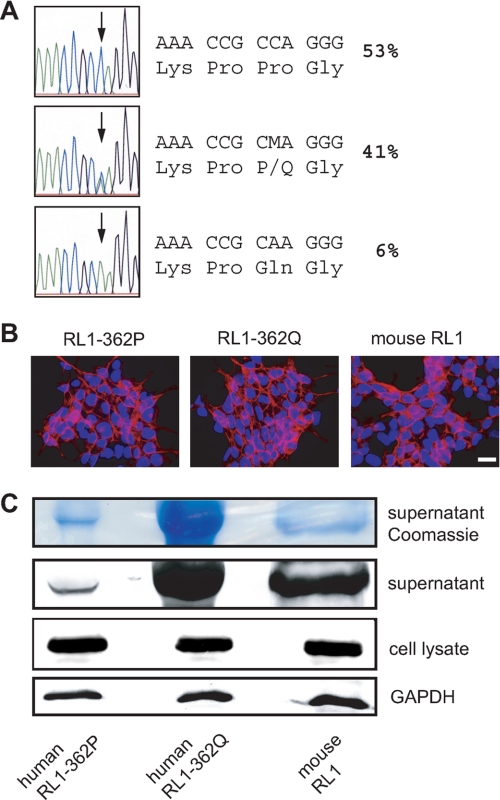
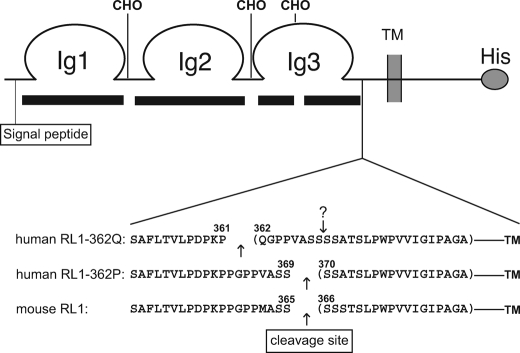
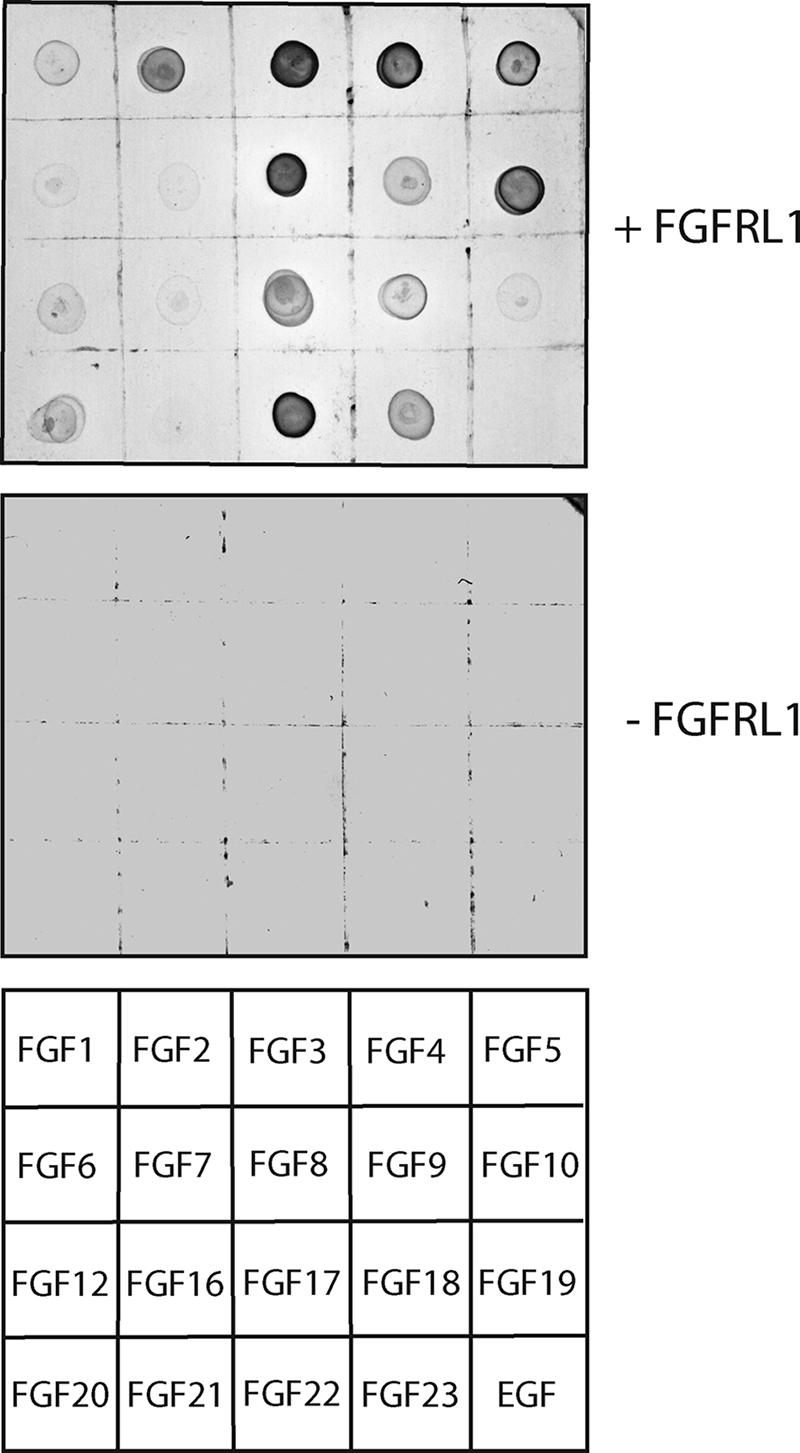
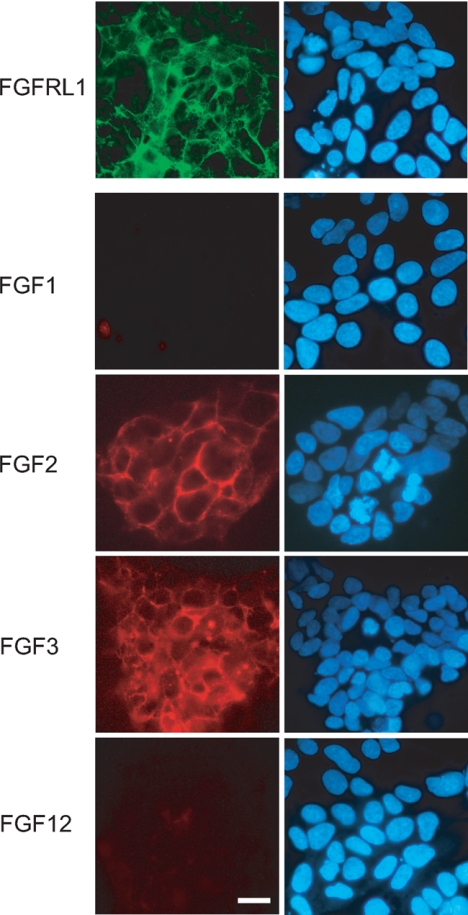
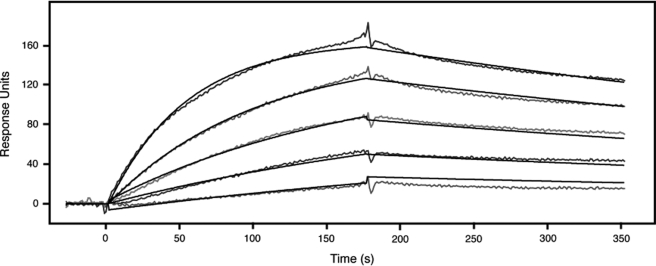
FGFRL1 (fibroblast growth factor receptor like 1) is the fifth and most recently discovered member of the fibroblast growth factor receptor (FGFR) family. With up to 50% amino acid similarity, its extracellular domain closely resembles that of the four conventional FGFRs. Its intracellular domain, however, lacks the split tyrosine kinase domain needed for FGF-mediated signal transduction. During embryogenesis of the mouse, FGFRL1 is essential for the development of parts of the skeleton, the diaphragm muscle, the heart, and the metanephric kidney. Since its discovery, it has been hypothesized that FGFRL1 might act as a decoy receptor for FGF ligands. Here we present several lines of evidence that support this notion. We demonstrate that the FGFRL1 ectodomain is shed from the cell membrane of differentiating C2C12 myoblasts and from HEK293 cells by an as yet unidentified protease, which cuts the receptor in the membrane-proximal region. As determined by ligand dot blot analysis, cell-based binding assays, and surface plasmon resonance analysis, the soluble FGFRL1 ectodomain as well as the membrane-bound receptor are capable of binding to some FGF ligands with high affinity, including FGF2, FGF3, FGF4, FGF8, FGF10, and FGF22. We furthermore show that ectopic expression of FGFRL1 in Xenopus embryos antagonizes FGFR signaling during early development. Taken together, our data provide strong evidence that FGFRL1 is indeed a decoy receptor for FGFs.
Figures

References
-
- Itoh N., Ornitz D. M. (2004) Trends Genet. 20, 563–569 - PubMed
-
- Itoh N., Ornitz D. M. (2008) Dev. Dyn 237, 18–27 - PubMed
-
- Wiedemann M., Trueb B. (2000) Genomics 69, 275–279 - PubMed
-
- Sleeman M., Fraser J., McDonald M., Yuan S., White D., Grandison P., Kumble K., Watson J. D., Murison J. G. (2001) Gene 271, 171–182 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
