

Differential requirements of MHC and of DCs for endogenous proliferation of different T-cell subsets in vivo
- PMID: 19920180
- PMCID: PMC2787171
- DOI: 10.1073/pnas.0909954106
Differential requirements of MHC and of DCs for endogenous proliferation of different T-cell subsets in vivo
Abstract
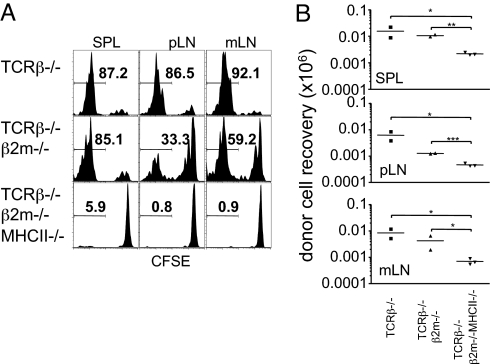
T cells transferred into severe lymphopenic hosts undergo rapid proliferation known as "endogenous proliferation" that are distinct from conventional homeostatic proliferation. Unlike homeostatic proliferation, cytokines, such as IL-7 are dispensable, yet TCR:MHC interaction is essential for this process to occur. However, cell types inducing the proliferation have not formally been addressed. In this study, we report that CD11c+ conventional DCs play irreplaceable roles in inducing endogenous proliferation of both naive and memory phenotype CD4 T cells via TCR-MHC II interaction. By contrast, CD8 T-cell endogenous proliferation was independent of MHC I or CD11c+ DCs. Interestingly, MHC II was necessary to support naive CD8 T-cell proliferation within MHC I-deficient hosts. Depletion of both B cells and DCs was sufficient to abrogate the proliferation of naive but not of memory CD8 T cells. These results suggest that depending on the T-cell lineages, as well as the differentiation status, different mechanisms control endogenous proliferation, revealing in vivo complexity of T-cell proliferation under lymphopenic conditions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Kieper WC, et al. Recent immune status determines the source of antigens that drive homeostatic T cell expansion. J Immunol. 2005;174:3158–3163. - PubMed
-
- Min B, Yamane H, Hu-Li J, Paul WE. Spontaneous and homeostatic proliferation of CD4 T cells are regulated by different mechanisms. J Immunol. 2005;174:6039–6044. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
