

Oncoviruses and Pathogenic MicroRNAs in Humans
- PMID: 19920887
- PMCID: PMC2778015
- DOI: 10.2174/1874357900903010037
Oncoviruses and Pathogenic MicroRNAs in Humans
Abstract
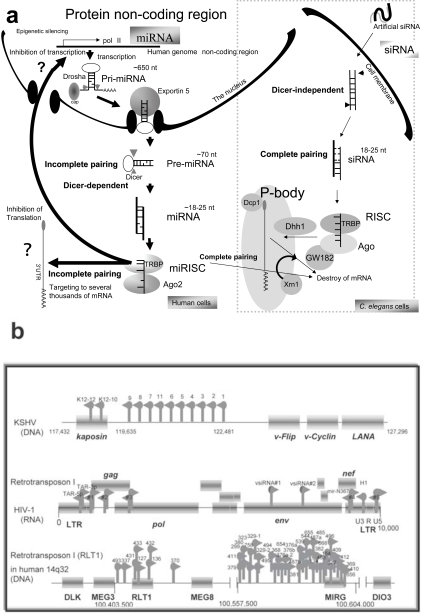
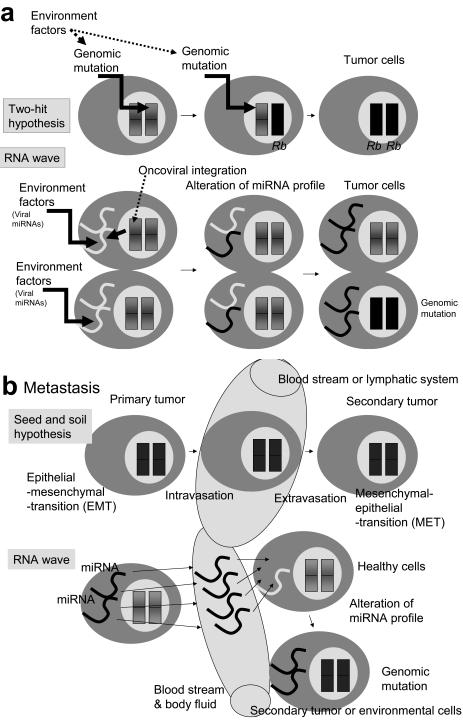
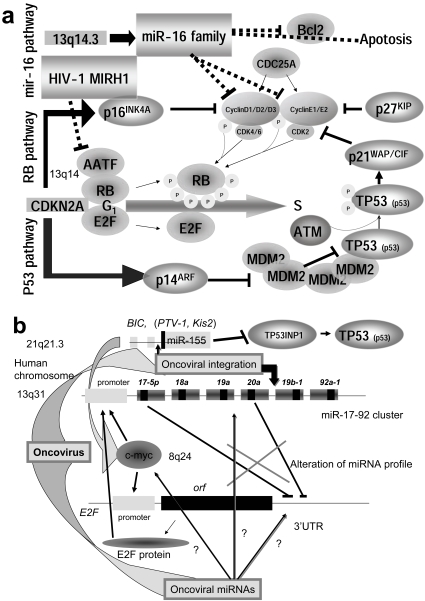
For disease prognosis, the functional significance of the oncoviral integration locus in oncogenesis has remained enigmatic. The locus encodes several transcripts without protein products, but microRNAs (miRNAs) have recently been identified from a common oncoviral integration locus. miRNA is an endogenous, non-coding small RNA by which gene expression is suppressed. Although miRNA genes, such as let-7 in the nematode, have orthologs among animals, the relationship between miRNAs and tumorigenesis or tumor suppression has been mainly discovered in several human cancers. On the contrary, this review clearly demonstrates the potential for human tumorigenesis of both miRNA genes from oncoviral integration sites and other cellular onco-microRNA genes, and we conclude that alteration of the miRNA profile of cells can be defined as tumorigenic or tumor suppressive. Thus, we explain here that virally-pathogenic miRNAs could also be partly responsible for oncogenesis or oncogene suppression to confirm' the RNA wave', with the miRNAs hypothesized as a mobile and functional genetic element.
Keywords: Cancer; human genome; microRNA; non-coding RNA; oncovirus; tumor..
Figures
Similar articles
-
A precursor microRNA in a cancer cell nucleus: get me out of here!Cell Cycle. 2011 Mar 15;10(6):922-5. doi: 10.4161/cc.10.6.15119. Epub 2011 Mar 15. Cell Cycle. 2011. PMID: 21346411 Free PMC article.
-
Emerging roles for natural microRNA sponges.Curr Biol. 2010 Oct 12;20(19):R858-61. doi: 10.1016/j.cub.2010.08.052. Curr Biol. 2010. PMID: 20937476 Free PMC article. Review.
-
Aberrant regulation of CXCR4 in cancer via deviant microRNA-targeted interactions.Epigenetics. 2022 Dec;17(13):2318-2331. doi: 10.1080/15592294.2022.2118947. Epub 2022 Sep 6. Epigenetics. 2022. PMID: 36047714 Free PMC article. Review.
-
MicroRNAs: non coding pleiotropic factors in development, cancer prevention and treatment.Microrna. 2013;2(2):81. doi: 10.2174/2211536611302020001. Microrna. 2013. PMID: 25070777
-
The microRNA expression signature of bladder cancer by deep sequencing: the functional significance of the miR-195/497 cluster.PLoS One. 2014 Feb 10;9(2):e84311. doi: 10.1371/journal.pone.0084311. eCollection 2014. PLoS One. 2014. PMID: 24520312 Free PMC article.
Cited by
-
Human Ribosomal RNA-Derived Resident MicroRNAs as the Transmitter of Information upon the Cytoplasmic Cancer Stress.Biomed Res Int. 2016;2016:7562085. doi: 10.1155/2016/7562085. Epub 2016 Jul 19. Biomed Res Int. 2016. PMID: 27517048 Free PMC article.
-
RNA Genes: Retroelements and Virally Retroposable microRNAs in Human Embryonic Stem Cells.Open Virol J. 2010 May 25;4:63-75. doi: 10.2174/1874357901004010063. Open Virol J. 2010. PMID: 20835360 Free PMC article.
-
MiR-26a promotes ovarian cancer proliferation and tumorigenesis.PLoS One. 2014 Jan 22;9(1):e86871. doi: 10.1371/journal.pone.0086871. eCollection 2014. PLoS One. 2014. PMID: 24466274 Free PMC article.
-
Tumor microbiome metabolism: A game changer in cancer development and therapy.Front Oncol. 2022 Jul 25;12:933407. doi: 10.3389/fonc.2022.933407. eCollection 2022. Front Oncol. 2022. PMID: 35936744 Free PMC article. Review.
References
-
- Chissoe SL, Bodenteich A, Wang Y-F, et al. Sequence and analysis of the human ABL gene, the BCR gene, and regions involved in the Philadelphia chromosomal translocation. Genomics. 1995;27:67–82. - PubMed
-
- Huppi K, Siwarski D. Chimeric transcripts with an open reading frame are generated as a result of translocation to the PVT-1 region in mouse B-cell tumors. Int J Cancer. 1994;59:848–51. - PubMed
-
- Palumbo AP, Boccadoro M, Battaglio S, et al. Human homologue of moloney leukemia virus integration-4 locus (MLVI-4), located 20 kilobases 3’ of the myc gene, is rearranged in multiple myelomas. Cancer Res. 1990;50:6478–82. - PubMed
LinkOut - more resources
Full Text Sources