

The endoplasmic reticulum chaperone Cosmc directly promotes in vitro folding of T-synthase
- PMID: 19923218
- PMCID: PMC2807302
- DOI: 10.1074/jbc.M109.065169
The endoplasmic reticulum chaperone Cosmc directly promotes in vitro folding of T-synthase
Abstract
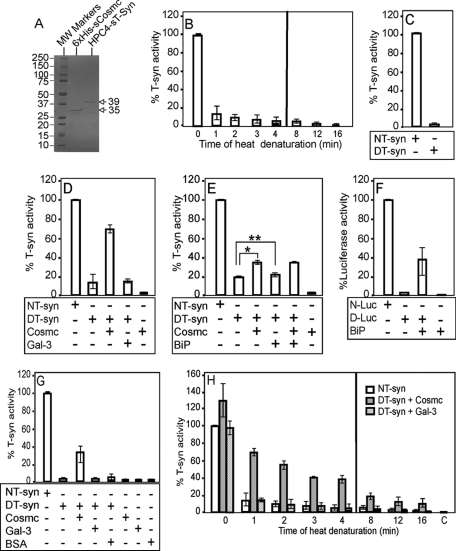
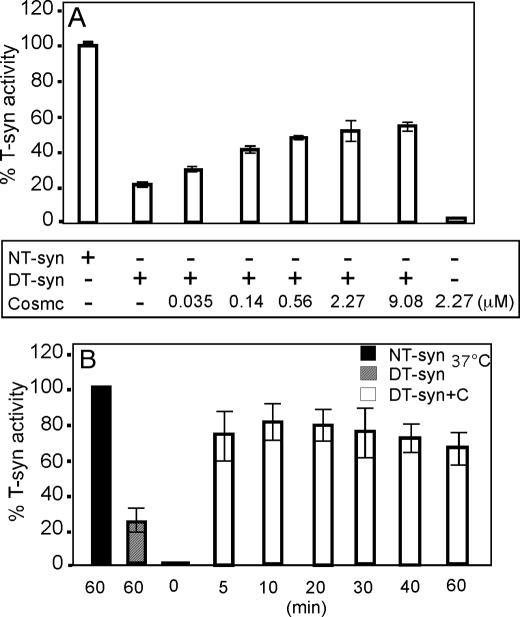
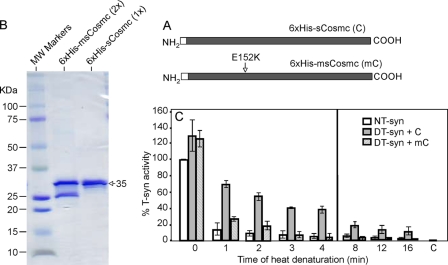
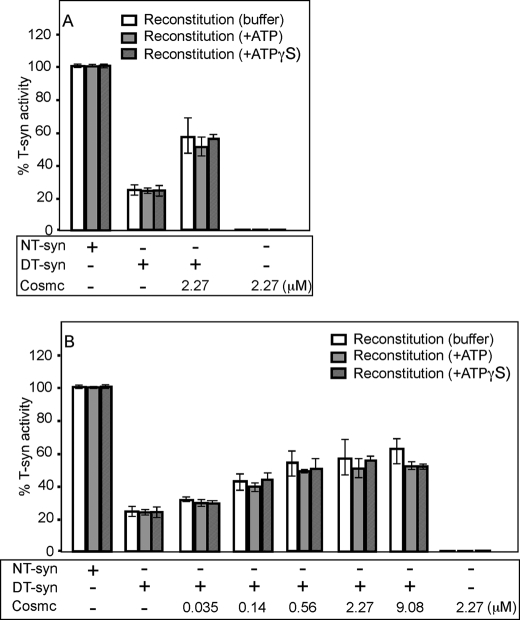
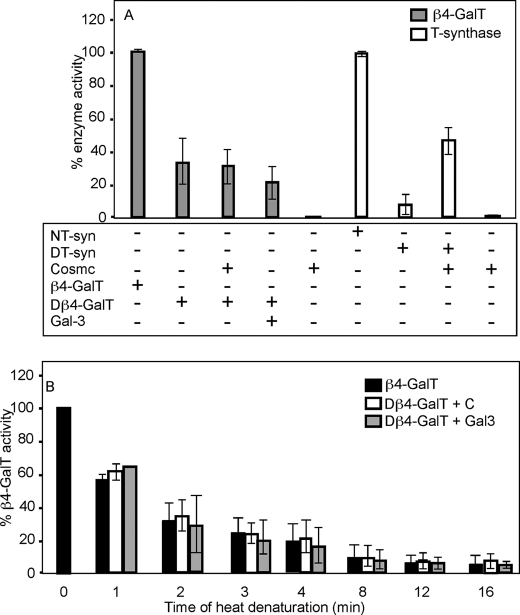
The T-synthase is the key beta 3-galactosyltransferase essential for biosynthesis of core 1 O-glycans (Gal beta 1-3GalNAc alpha 1-Ser/Thr) in animal cell glycoproteins. Here we describe the novel ability of an endoplasmic reticulum-localized molecular chaperone termed Cosmc to specifically interact with partly denatured T-synthase in vitro to cause partial restoration of activity. By contrast, a mutated form of Cosmc observed in patients with Tn syndrome has reduced chaperone function. The chaperone activity of Cosmc is specific, does not require ATP in vitro, and is effective toward T-synthase but not another beta-galactosyltransferase. Cosmc represents the first ER chaperone identified to be required for folding of a glycosyltransferase.
Figures
References
-
- Helenius A., Marquardt T., Braakman I. (1992) Trends Cell Biol. 2, 227–231 - PubMed
-
- Hebert D. N., Molinari M. (2007) Physiol. Rev. 87, 1377–1408 - PubMed
-
- Ellgaard L., Helenius A. (2003) Nat. Rev. Mol. Cell Biol. 4, 181–191 - PubMed
-
- Anfinsen C. B. (1973) Science 181, 223–230 - PubMed
-
- Fink A. L. (1999) Physiol. Rev. 79, 425–449 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
