

Techniques and tactics used in determining the structure of the trimeric ebolavirus glycoprotein
- PMID: 19923712
- PMCID: PMC2777170
- DOI: 10.1107/S0907444909032314
Techniques and tactics used in determining the structure of the trimeric ebolavirus glycoprotein
Abstract
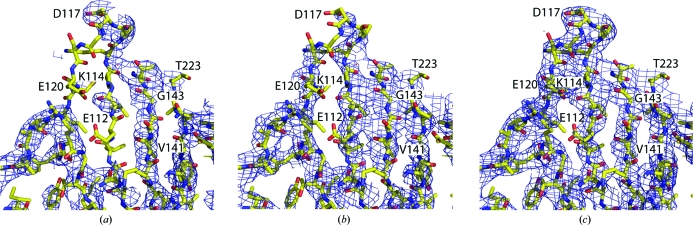
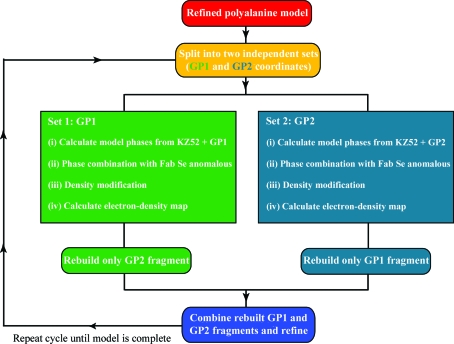
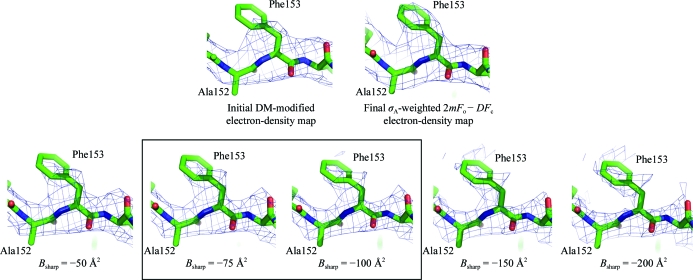
The trimeric membrane-anchored ebolavirus envelope glycoprotein (GP) is responsible for viral attachment, fusion and entry. Knowledge of its structure is important both for understanding ebolavirus entry and for the development of medical interventions. Crystal structures of viral glycoproteins, especially those in their metastable prefusion oligomeric states, can be difficult to achieve given the challenges in production, purification, crystallization and diffraction that are inherent in the heavily glycosylated flexible nature of these types of proteins. The crystal structure of ebolavirus GP in its trimeric prefusion conformation in complex with a human antibody derived from a survivor of the 1995 Kikwit outbreak has now been determined [Lee et al. (2008), Nature (London), 454, 177-182]. Here, the techniques, tactics and strategies used to overcome a series of technical roadblocks in crystallization and phasing are described. Glycoproteins were produced in human embryonic kidney 293T cells, which allowed rapid screening of constructs and expression of protein in milligram quantities. Complexes of GP with an antibody fragment (Fab) promoted crystallization and a series of deglycosylation strategies, including sugar mutants, enzymatic deglycosylation, insect-cell expression and glycan anabolic pathway inhibitors, were attempted to improve the weakly diffracting glycoprotein crystals. The signal-to-noise ratio of the search model for molecular replacement was improved by determining the structure of the uncomplexed Fab. Phase combination with Fab model phases and a selenium anomalous signal, followed by NCS-averaged density modification, resulted in a clear interpretable electron-density map. Model building was assisted by the use of B-value-sharpened electron-density maps and the proper sequence register was confirmed by building alternate sequences using N-linked glycan sites as anchors and secondary-structural predictions.
Figures

Similar articles
-
Structure of the Ebola virus glycoprotein bound to an antibody from a human survivor.Nature. 2008 Jul 10;454(7201):177-82. doi: 10.1038/nature07082. Nature. 2008. PMID: 18615077 Free PMC article.
-
Biochemical and structural characterization of cathepsin L-processed Ebola virus glycoprotein: implications for viral entry and immunogenicity.J Virol. 2010 Mar;84(6):2972-82. doi: 10.1128/JVI.02151-09. Epub 2010 Jan 6. J Virol. 2010. PMID: 20053739 Free PMC article.
-
The Tetherin Antagonism of the Ebola Virus Glycoprotein Requires an Intact Receptor-Binding Domain and Can Be Blocked by GP1-Specific Antibodies.J Virol. 2016 Nov 28;90(24):11075-11086. doi: 10.1128/JVI.01563-16. Print 2016 Dec 15. J Virol. 2016. PMID: 27707924 Free PMC article.
-
Neutralizing ebolavirus: structural insights into the envelope glycoprotein and antibodies targeted against it.Curr Opin Struct Biol. 2009 Aug;19(4):408-17. doi: 10.1016/j.sbi.2009.05.004. Epub 2009 Jun 24. Curr Opin Struct Biol. 2009. PMID: 19559599 Free PMC article. Review.
-
Structural basis for differential neutralization of ebolaviruses.Viruses. 2012 Apr;4(4):447-70. doi: 10.3390/v4040447. Epub 2012 Apr 5. Viruses. 2012. PMID: 22590681 Free PMC article. Review.
Cited by
-
Structures of protective antibodies reveal sites of vulnerability on Ebola virus.Proc Natl Acad Sci U S A. 2014 Dec 2;111(48):17182-7. doi: 10.1073/pnas.1414164111. Epub 2014 Nov 17. Proc Natl Acad Sci U S A. 2014. PMID: 25404321 Free PMC article.
-
A convenient and general expression platform for the production of secreted proteins from human cells.J Vis Exp. 2012 Jul 31;(65):4041. doi: 10.3791/4041. J Vis Exp. 2012. PMID: 22872008 Free PMC article.
-
Enhancement of Ebola Virus Infection via Ficolin-1 Interaction with the Mucin Domain of GP Glycoprotein.J Virol. 2016 May 12;90(11):5256-5269. doi: 10.1128/JVI.00232-16. Print 2016 Jun 1. J Virol. 2016. PMID: 26984723 Free PMC article.
-
Structural Biology Illuminates Molecular Determinants of Broad Ebolavirus Neutralization by Human Antibodies for Pan-Ebolavirus Therapeutic Development.Front Immunol. 2022 Jan 10;12:808047. doi: 10.3389/fimmu.2021.808047. eCollection 2021. Front Immunol. 2022. PMID: 35082794 Free PMC article. Review.
-
A shared structural solution for neutralizing ebolaviruses.Nat Struct Mol Biol. 2011 Nov 20;18(12):1424-7. doi: 10.1038/nsmb.2150. Nat Struct Mol Biol. 2011. PMID: 22101933 Free PMC article.
References
-
- Abrahams, J. P. & Leslie, A. G. W. (1996). Acta Cryst. D52, 30–42. - PubMed
-
- Adams, P. D., Gopal, K., Grosse-Kunstleve, R. W., Hung, L.-W., Ioerger, T. R., McCoy, A. J., Moriarty, N. W., Pai, R. K., Read, R. J., Romo, T. D., Sacchettini, J. C., Sauter, N. K., Storoni, L. C. & Terwilliger, T. C. (2004). J. Synchrotron Rad.11, 53–55. - PubMed
-
- Adams, P. D., Grosse-Kunstleve, R. W., Hung, L.-W., Ioerger, T. R., McCoy, A. J., Moriarty, N. W., Read, R. J., Sacchettini, J. C., Sauter, N. K. & Terwilliger, T. C. (2002). Acta Cryst. D58, 1948–1954. - PubMed
-
- Afonine, P. D., Grosse-Kunstleve, R. W. & Adams, P. D. (2005). CCP4 Newsl., contribution 8.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
